Содержащиеся в цитоплазме неоднородные по форме митохондрии и микросомы размером около 0,1...5 мкм, то есть собственно центры ферментативных преобразований при обмене веществ, в противоположность клеточным перегородкам и наружной оболочке цитоплазмы сохраняют свою видимую структуру до послеклимактерического периода [255]. Изменения в составе жирных кислот митохондрий и микросом позволяют сделать вывод о том, что в них также происходят очень тонкие структурные изменения мембран. Особенно под действием этилена [159] митохондрии увеличиваются в объеме и проницаемость мембран возрастает. Этим отчасти можно объяснить их повышенную активность обмена веществ и возрастающую интенсивность дыхания при созревании плодов.[ ...]
Все эти процессы в значительной степени зависят от температуры и с ее повышением в камере хранилища протекают быстрее. Они замедляются при низком содержании кислорода и высокой концентрации двуокиси углерода в газовой среде камеры. Под их влиянием структура клетки сохраняется дольше и клетки не теряют тургора. Поэтому плоды, хранившиеся в регулируемой газовой среде, ко времени выгрузки их из хранилища отличаются более плотной консистенцией и меньшей степенью зрелости.[ ...]
Диссимиляция углеводов может происходить двумя путями. В плодах семечковых пород сахар в основном расходуется на дыхание по пути ЭМ.П (Эмбдена — Мей-ерхофа — Парнаса) [307, 308]. При этом в связи с процессами фосфорилирования глюкоза расщепляется до пировиноградиой кислоты (гликолиз). Кроме того, существует возможность расщепления углеводов по пентоз-ному циклу [89]. В каком размере этот цикл участвует в преобразовании дыхательных субстратов, пока нельзя сказать. В настоящее время предполагается, что на определенных этапах развития яблока [88] или других плодов преобладает тот или иной путь. Преобладающий у плодов семечковых пород путь ЭМП заканчивается пировиноградиой кислотой, которая играет важнейшую роль в дыхании. С этого момента дальнейшие превращения пировиноградиой кислоты зависят от среды: в аэробных — с расходованием кислорода, в анаэробных — когда кислород не требуется.[ ...]
Образование энергии без кислорода протекает в соответствии с уравнением СеН Ое—у2С2Н5ОН + 2СО2Н-+ 28,2 ккал. Этот путь обозначают как «брожение» или «анаэробное дыхание». В клетках в качестве побочных продуктов появляются альдегид уксусной кислоты, уксусная кислота, ацетоин, диацетил и другие соединения, которые частично являются компонентами летучих веществ, выделяемых плодами при их созревании.[ ...]
Как видно из уравнений, при обоих типах дыхания освобождаются разные количества энергии. Так как клетка стремится возможно рациональнее регулировать преобразование веществ, аэробное дыхание играет намного большую роль, чем анаэробное, тем более что при первом, как правило, теряется меньше сахаров.[ ...]
Куиревич [149] установил критическую концентрацию равной 10%, однако Том кине [288] считает, что она должна быть ниже 5% (¡рис. 63). Из этого необходимо сделать вывод, что диапазон, в котором аэробное дыхание и брожение протекают одновременно, определяется сортовыми и породными особенностями плодов и сильно колеблется в зависимости от условий вегетации и агротехники. Возможно, этим объясняется причина различного поведения видов и сортов плодов в отдельные годы их хранения в камерах с регулируемой газовой средой.[ ...]
Поскольку, по Ульриху (1957), критическая концентрация кислорода у яблок в каждом случае достигает менее 3% в холодильных камерах с регулируемой газовой средой, ее лишь в исключительных случаях следует снижать ниже этого уровня.[ ...]
При сравнении обоих типов дыхания бросается в глаза неодинаковое отношение поглощения кислорода к выделению двуокиси углерода. Отношение СО2/О2 обозначается как дыхательный коэффициент КО.[ ...]
Дыхательный коэффициент при нормальном созревании плодов не остается постоянным. В предклимакте-рической стадии он равен примерно 1 и по мере созревания достигает значений 1,2... 1,5. При отклонениях на ±0,25 от единицы в плодах еще не наблюдаются аномалии в обмене веществ, и только при больших отклонениях можно предполагать физиологические расстройства. Интенсивность дыхания отдельных слоев тканей какого-либо плода неодинакова. В соответствии с большей активностью ферментов в кожице в ней отмечается во много раз большая интенсивность дыхания, чем у паренхимной ткани (Хулме и Родс, 1939). При снижении содержания кислорода и увеличении концентрации двуокиси углерода в паренхимных клетках по мере удаления от кожицы к сердцевине плода интенсивность дыхания падает.[ ...]
Местом ферментативных преобразований и, следовательно, биологического окисления считаются органеллы протоплазмы, и прежде всего митохондрии [18, Хултин, 1959]. Поэтому их структурные и химические изменения в ходе созревания плодов, по данным Хоарда [105], заметны по уровню интенсивности дыхания.[ ...]
Рисунки к данной главе:
| Влияние концентрации кислорода на интенсивность дыхания плодов [149] (интенсивность дыхания в обычном воздухе = = 100%) |
![Влияние концентрации кислорода на интенсивность дыхания плодов [149] (интенсивность дыхания в обычном воздухе = = 100%)](/static/pngsmall/826585254.png) |
| Интенсивность дыхания в процессе развития плодов яблони (по Шульцу, 1970 г.) |
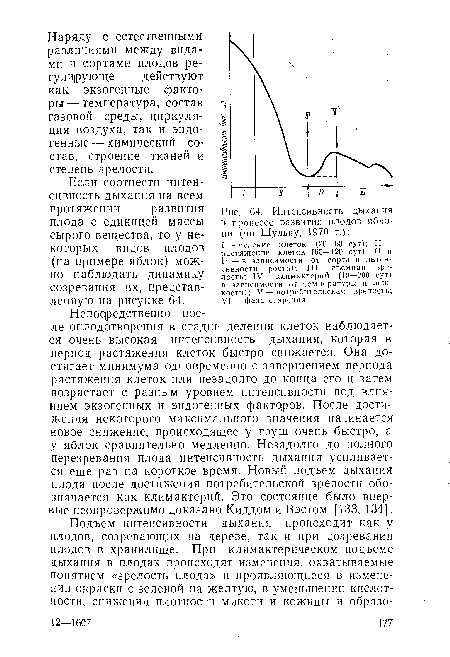 |
| Влияние температуры на интенсивность ды-хания груш сорта Вильямс [136]. I |
![Влияние температуры на интенсивность ды-хания груш сорта Вильямс [136]. I](/static/pngsmall/826585264.png) |
| Влияние температуры хранения на леж-кость яблок и груш [147] |
![Влияние температуры хранения на леж-кость яблок и груш [147]](/static/pngsmall/826585266.png) |
Аналогичные главы в дргуих документах:
| См. далее:Дыхание |
| См. далее:Дыхание |
| См. далее:Дыхание |