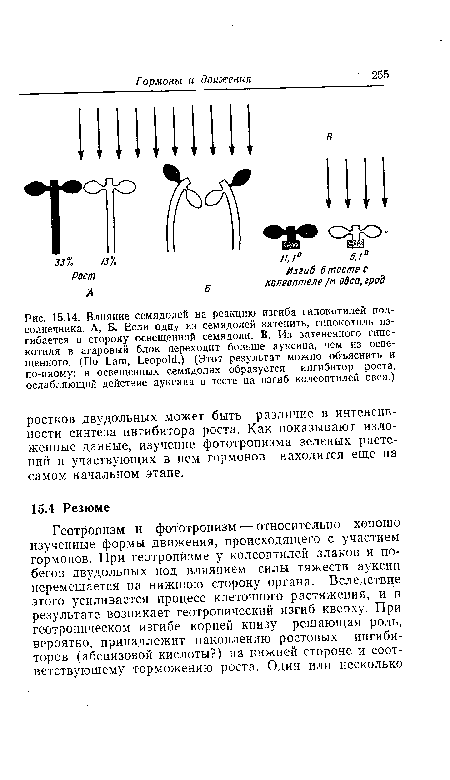
Фототропический изгиб, так же как и геотропический, обусловлен различием в скорости роста освещенной ;1 неосвещенной сторон растущего органа. В случае положительного фототропизма, который обычно свойствен побегам при естественных уровнях освещенности, теневая сторона растет быстрее, чем обращенная к свету. Так же как при геотропизме, изгиб начинается от верхушки органа и распространяется базипетально, а выше изгиба вновь происходит распрямление (см. рис. 15.8). Изучать фототропический изгиб значительно труднее, чем геотропический. В то время как сила тяжести практически постоянна, действие света обнаруживает характерную зависимость от длины волны и дозы. Мы не будем рассматривать спектр действия и связанный с этим вопрос о фоторецепторе, так как нет сомнений в том, что гормоны в стадии восприятия фототропического раздражения не участвуют.[ ...]
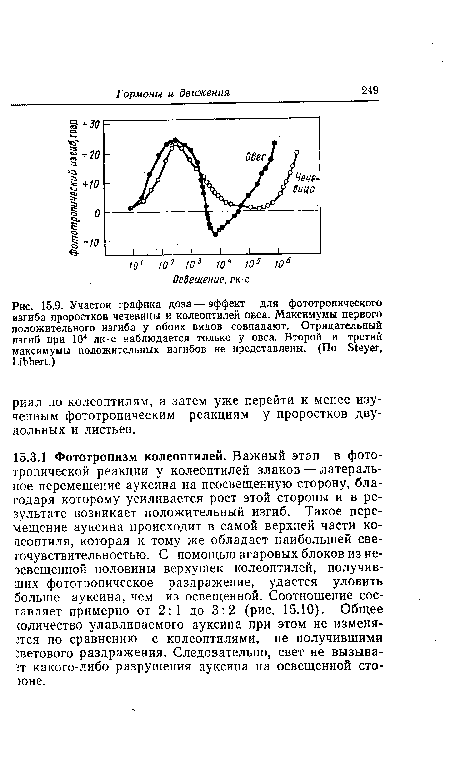
График доза—эффект для фототропизма имеет вид многовершинной кривой. При увеличении дозы (произведение интенсивности освещения на время) выявляются два максимума реакции, а при особенно высоких интенсивностях света и его непрерывном воздействии намечается даже подъем к третьему максимуму. Между областями максимальной эффективности располагаются два минимума. У колеоптилей овса первый минимум лежит в отрицательной области, т. е. при соответствующих дозах (около 104 лк-с) колеоптили изгибаются по направлению от света. Максимумы обозначают как 1-й, 2-й и иногда 3-й положительные изгибы, а лежащий в отрицательной области минимум — как 1-й отрицательный изгиб (рис. 15.9). Важнейшие гипотезы о природе фототропизма основаны на данных, полученных в области 1-го положительного изгиба. В природных условиях получаемые растением количества света чаще всего соответствуют области 2-го и 3-го положительных изгибов. Разумеется, при изучении фототропизма очень важно учитывать зависимость эффекта от дозы.[ ...]
Другая трудность при исследовании явления фототропизма связана с тем, что каждый фототропический изгиб немедленно вызывает встречную геотропическую реакцию. Геотропизм, напротив, можно изучать без каких-либо затруднений в отсутствие фототропически действующего света.[ ...]
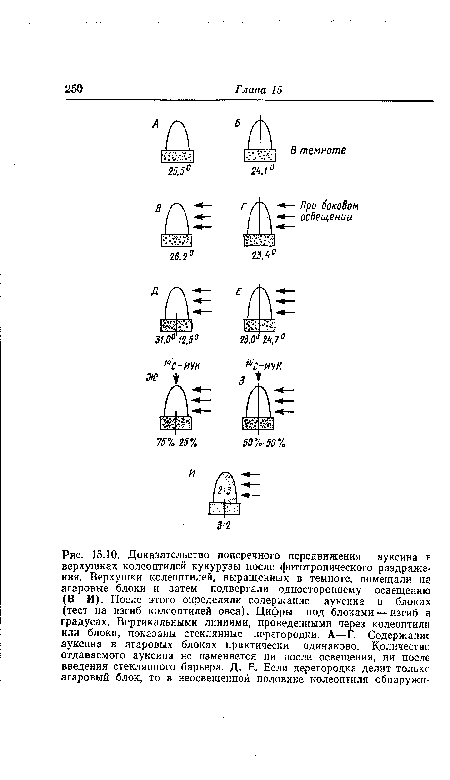
Этот факт поперечной поляризации тока ауксина был впервые установлен Вентом в 20-х годах нашего столетия, а позднее получил подтверждение в многочисленных экспериментах, в том числе с применением меченой ИУК (рис. 15.10). Повышенная отдача ауксина в агаровые блоки на неосвещенной стороне, вероятно, объясняется двумя причинами — торможением базипетального транспорта ауксина под влиянием света и стимуляцией светом латерального транспорта. Пока еще не ясно, которое из этих двух изменений происходит раньше и зависимы ли они друг от друга. Доказано торможение транспорта ауксина фототропно-эффективным синим светом, и можно себе представить, что такого рода торможение на освещенной стороне приводит к перетеканию ауксина на теневую сторону, т. е. к его латеральному переносу. Если под действием света прекращается базипетальный отток ауксина, то в освещенной стороне его теперь должно быть больше, чем в затененной, где он беспрепятственно транспортируется. Так оно. и есть в действительности (рис. 15.10, И).[ ...]
Поскольку у колеоптилей овса при определенных дозах света (см. рис. 15.9) наблюдается отрицательный фототропический изгиб, то в соответствии с гипотезой Вента следует ожидать, что в эксперименте с улавливанием ауксина, представленном на рис. 15.10, Д, в агаровом блоке на освещенной стороне окажется больше ауксина. Это предположение тоже подтвердилось.[ ...]
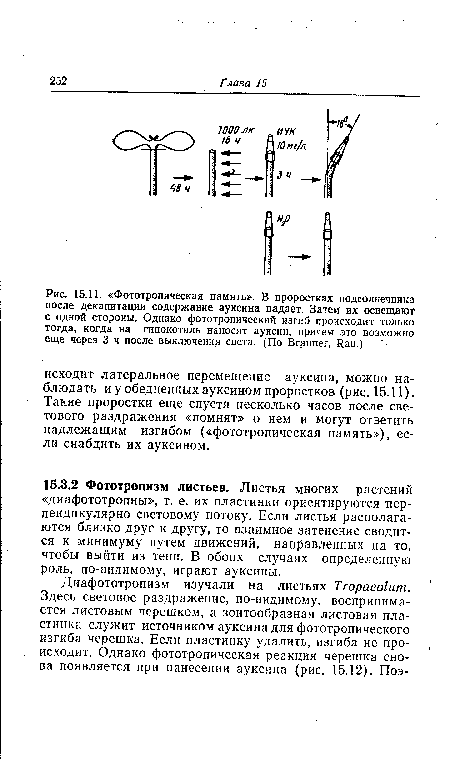
Природа этого движения, направленного на выход из тени, более подробно изучена на листьях Fatsia (рис. 15.13). В опытах с улавливанием ауксина в агаровые блоки было показано, что из затененной половины листовой пластинки в блок поступает больше активного ауксина, чем из освещенной. Очевидно, здесь в противоположность тому, что происходит в колеоптилях кукурузы (ср. рис. 15.10), образование ауксина или его транспорт стимулируется затенением. Можно, правда, предположить, что в затененной стороне образуется меньше ингибиторов роста или же заторможен их транспорт: результат был бы тот же — изгиб черешка в направлении освещенной половины листовой пластинки. Некоторые данные об участии ингибиторов роста в фототропизме были получены в опытах с проростками двудольных.[ ...]
Рисунки к данной главе:
| Фототропический изгиб колеоптиля овса после освещения (4 с, 30 лк). Изгиб начинается от верхушки и распространяется бази-петально, причем верхушка снова распрямляется. (По Апэг.) |
 |
| Если одну половину листа Лг^га затенить, то лист изгибается так, чтобы выйти из тени. Из затененной половины в агаровый блок переходит больше ауксина, чем из незатененной. (По Уашапе.) |
 |
Аналогичные главы в дргуих документах:
| См. далее:Фототропизм |