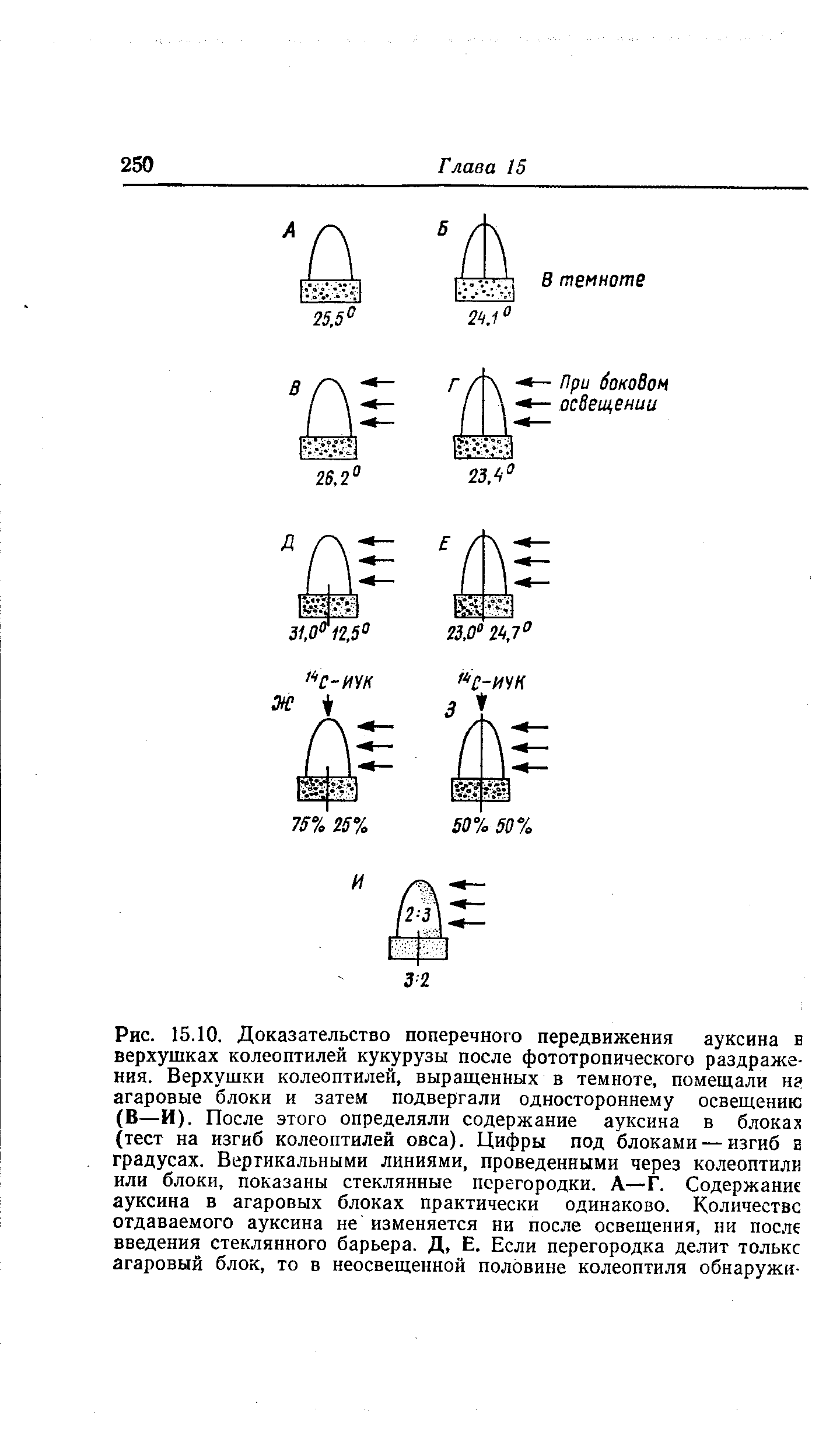
Этот факт поперечной поляризации тока ауксина был впервые установлен Вентом в 20-х годах нашего столетия, а позднее получил подтверждение в многочисленных экспериментах, в том числе с применением меченой ИУК (рис. 15.10). Повышенная отдача ауксина в агаровые блоки на неосвещенной стороне, вероятно, объясняется двумя причинами — торможением базипетального транспорта ауксина под влиянием света и стимуляцией светом латерального транспорта. Пока еще не ясно, которое из этих двух изменений происходит раньше и зависимы ли они друг от друга. Доказано торможение транспорта ауксина фототропно-эффективным синим светом, и можно себе представить, что такого рода торможение на освещенной стороне приводит к перетеканию ауксина на теневую сторону, т. е. к его латеральному переносу. Если под действием света прекращается базипетальный отток ауксина, то в освещенной стороне его теперь должно быть больше, чем в затененной, где он беспрепятственно транспортируется. Так оно. и есть в действительности (рис. 15.10, И).
Вернуться к оглавлению