| Различные стадии прорастания зерна пшеницы. (По ^тев.) |
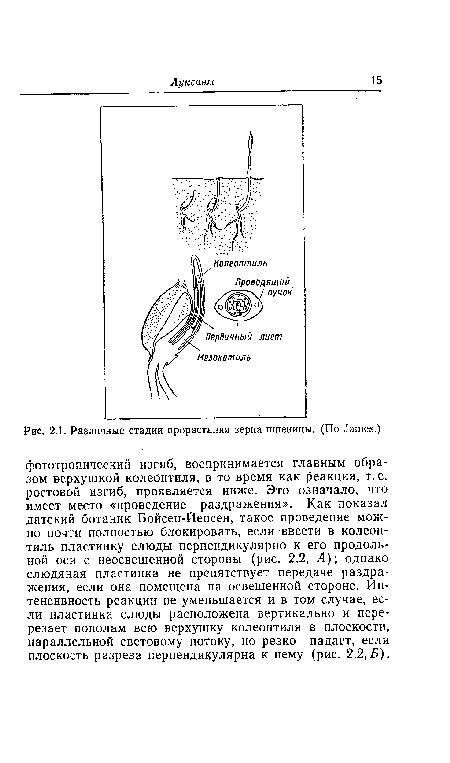 |
Далее
| Структура индолил-3-уксусной кислоты, которую называют также индол-3-ил-уксусной кислотой, (5-индолилуксусной кислотой или сокращенно ИУК.. |
 |
Далее
| Стимулирующее действие ИУК на клеточное деление в камбии молодых эпикотилей подсолнечника. А — в эпикотиль вводили ИУК путем нанесения ИУК-содержащей пасты на поверхность среза после удаления верхушки побега. Б — контроль (паста без ИУК) |
 |
Далее
| ИУК и апикальное доминирование. Де-капитированные проростки подсолнечника были обработаны пастой, не содержащей ауксина (А), и ланолиновой пастой с 1% ИУК (Б). ИУК подавляет развитие боковых побегов в пазухах семядолей. |
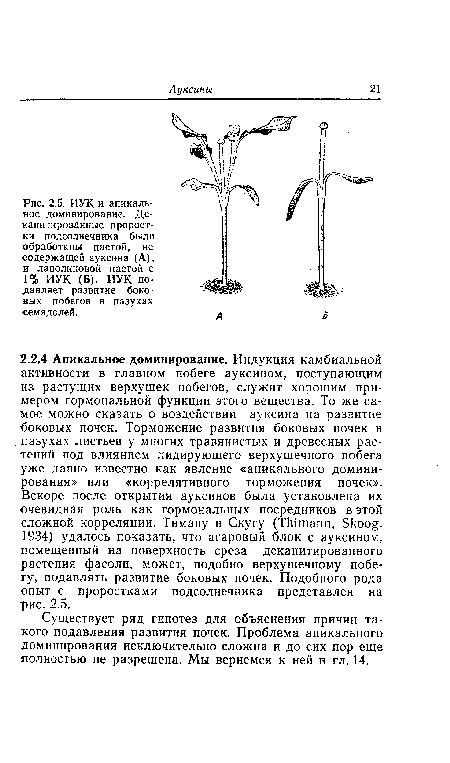 |
Далее
| Важнейшие природные индольные ауксины и возможные-пути биосинтеза индолил-3-уксусной кислоты. |
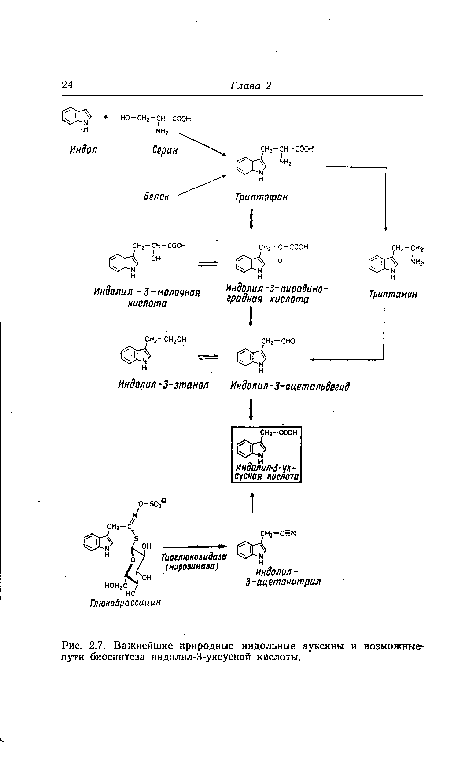 |
Далее
| Некоторые другие ауксины, в том числе не содержащие систему колец индола. |
 |
Далее
| Некоторые важные синтетические ауксины. |
 |
Далее
| Схема методических приемов для выделения, очистки и качественного и количественного определения ИУК из проростков гороха. Растительный экстракт (А) очишают методом тонкослойной хроматографии (Б, В), после чего проводят оценку биологической активности (Г, Д). Для этого используют отрезки колеоптилей, рост |
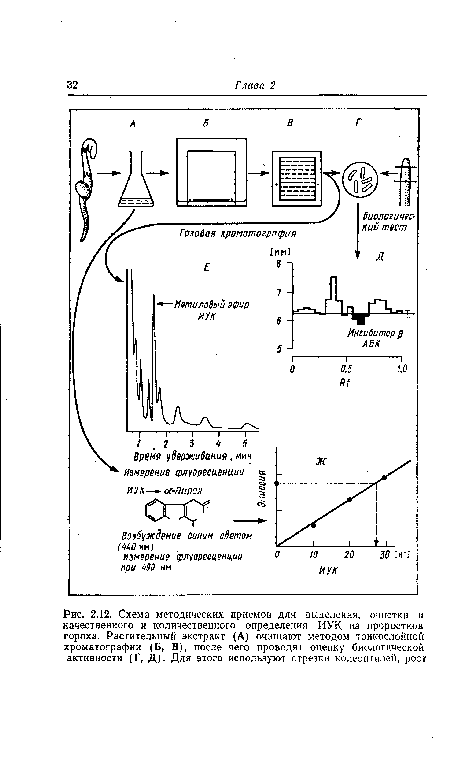 |
Далее
| График доза — эффект для ИУК. Работа проводилась с отрезками колеопти-лей овса длиной 5 мм. Ордината |
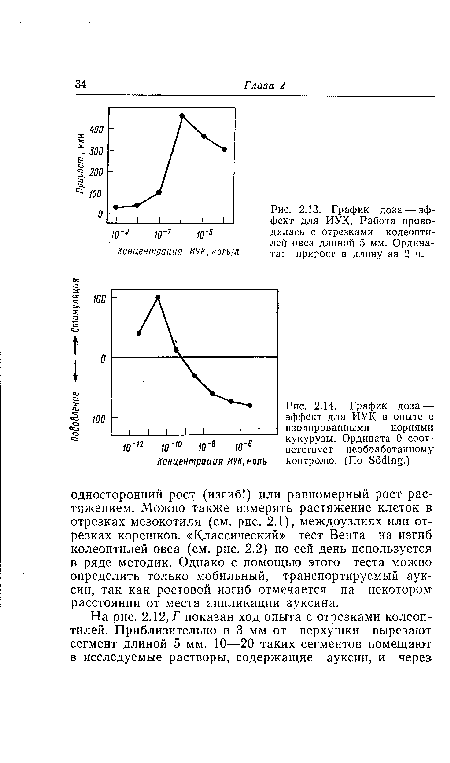 |
Далее
| График доза — эффект для ИУК в опыте с изолированными корнями кукурузы. Ордината 0 соответствует необработанному контролю. (По БосИпй.) |
|
Далее
| Основные этапы распада и инактивации ИУК. |
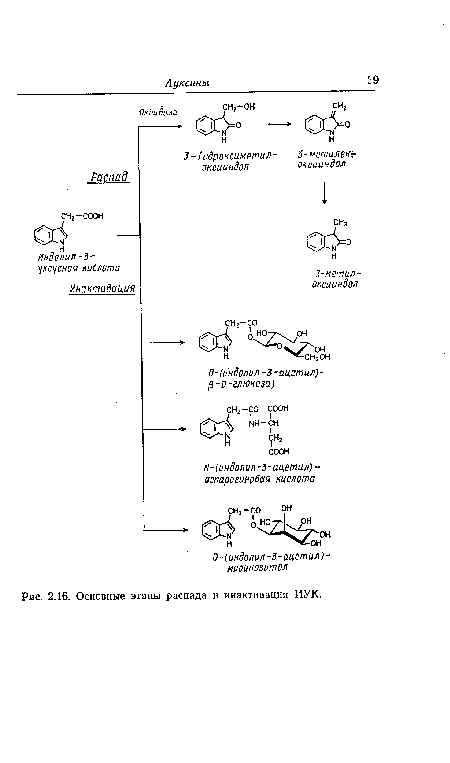 |
Далее
| Распределение активности ИУК-оксидазы, измеренной по скорости расщепления ИУК тканями (мкг ИУК/мг N в 1 ч), у проростка гороха. (По Galston, Dalberg.) |
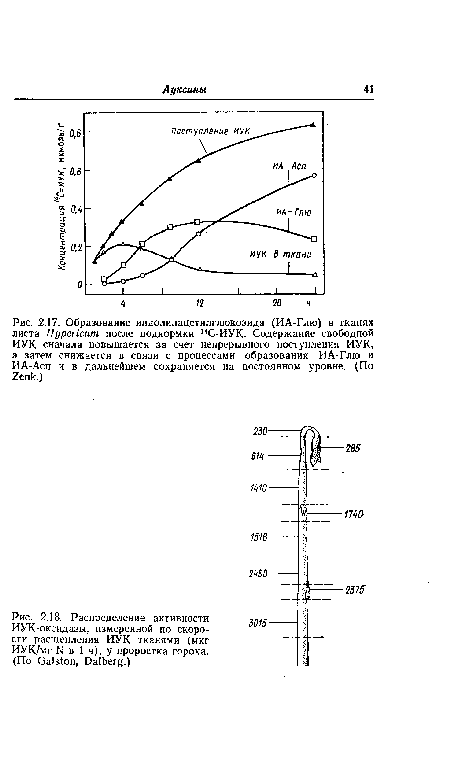 |
Далее
| Активность ауксина и вторичное утолщение камбия в ветвях клена. Количества ауксина представлены как углы изгиба в пробе с колеоптилями овса (см. рис. 2.2) в расчете на одинаковое количество ткани камбия. (По БбШщ .) |
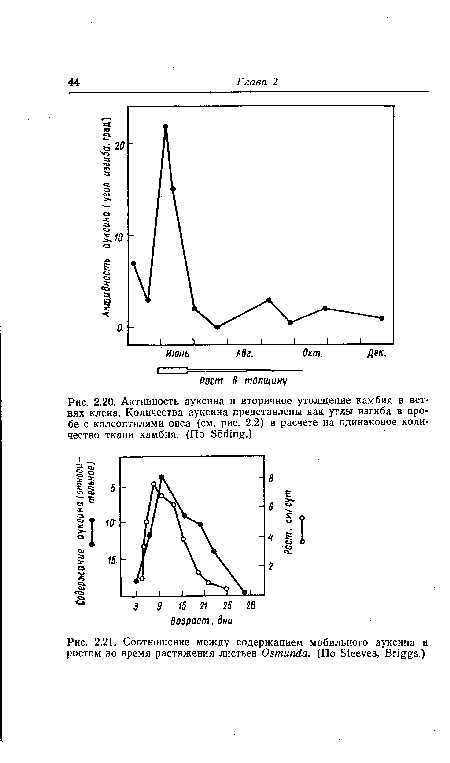 |
Далее
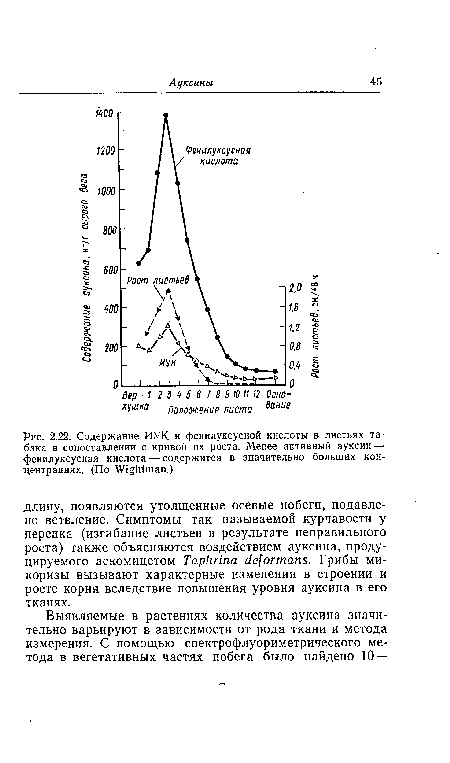
| Содержание ИУК и фенилуксусной кислоты в листьях табака в сопоставлении с кривой их роста. Менее активный ауксин — фенилуксусная кислота — содержится в значительно больших концентрациях. (По Wightman.) |
 |
Далее
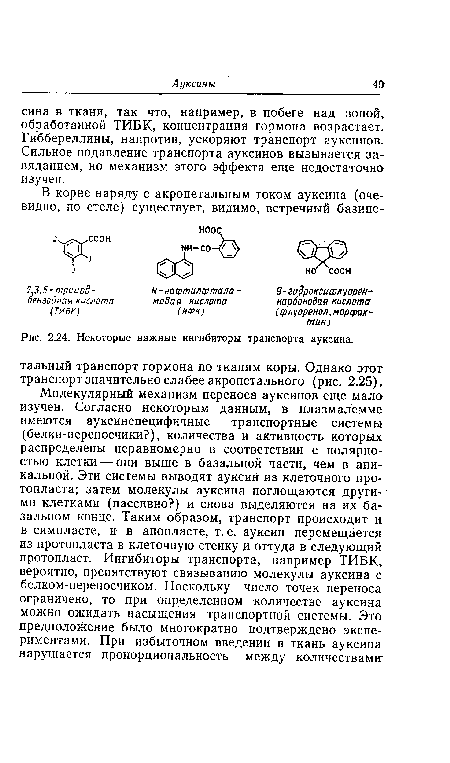
| Некоторые важные ингибиторы транспорта ауксина. |
 |
Далее
| Передвижение радиоактивной ИУК по отрезкам корней кукурузы длиной 6 мм. Транспорт в направлении к кончику корня (акропетальный, а) значительно лучше выражен, чем транспорт к основанию корня (базипетальный, б). По мере удаления от кончика корня интенсивность передвижения падает. (По Ва(ха.) |
 |
Далее
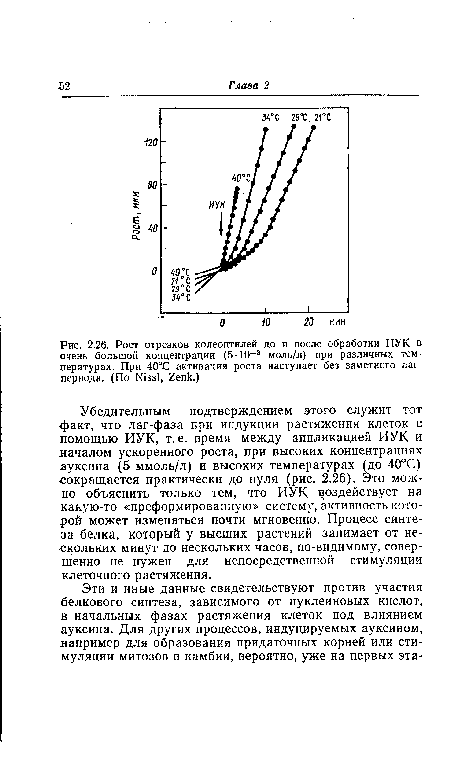
| Рост отрезков колеоптилей до и после обработки ИУК в очень большой концентрации (5-10-3 моль/л) при различных температурах. При 40°С активация роста наступает без заметного лаг-периода. (По Nissl, Zenk.) |
 |
Далее
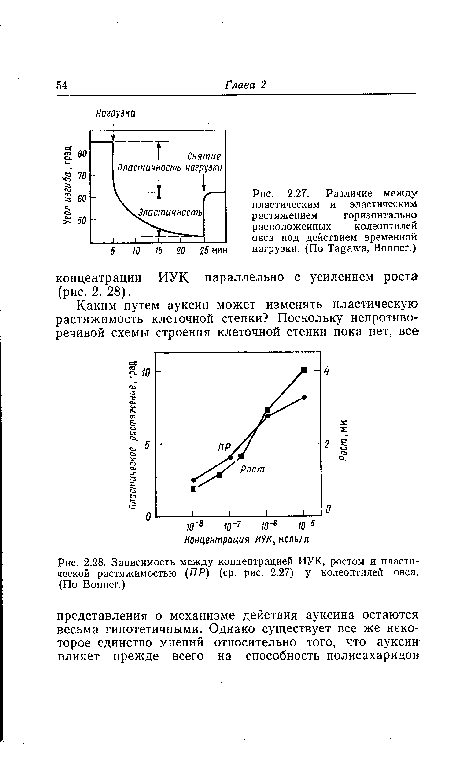
| Различие между пластическим и эластическим растяжением горизонтально расположенных колеоптилей овса под действием временной 5 10 15 20 25 мин нагрузки. (По Tagawa, Bonner.) |
 |
Далее
| Зависимость между концентрацией ИУК, ростом и пластической растяжимостью (ПР) (ср. рис. 2.27) у колеоптилей овса. (По Bonner.) |
|
Далее
| Соотношение между выбросом протонов и ростом отрезков колеоптилей. При повышении концентрации ИУК от 5-10 |
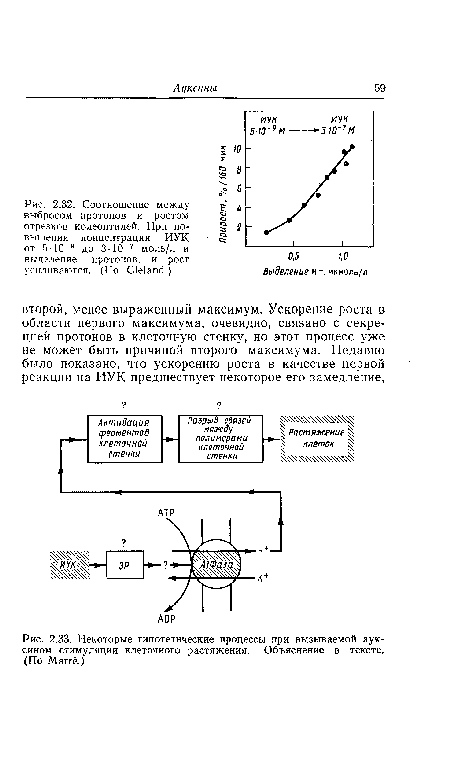 |
Далее
| Некоторые гипотетические процессы при вызываемой ауксином стимуляции клеточного растяжения. Объяснение в тексте. (По Marré.) |
|
Далее
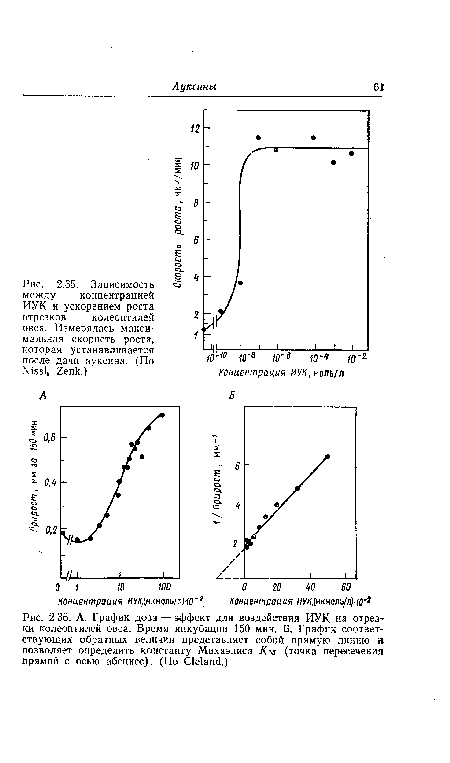
| Зависимость между концентрацией ИУК и ускорением роста отрезков колеоптилей овса. Измерялась максимальная скорость роста, которая устанавливается после дачи ауксина. (По N¡551» 2епк.) |
 |
Далее
| А. График доза — эффект для воздействия ИУК на отрезки колеоптилей овса. Время инкубации 150 мин. Б. График соответствующих обратных величин представляет собой прямую линию и позволяет определить константу Михаэлиса К м (точка пересечения прямой с осью абсцисс). (По С1е1ап(1.) |
|
Далее
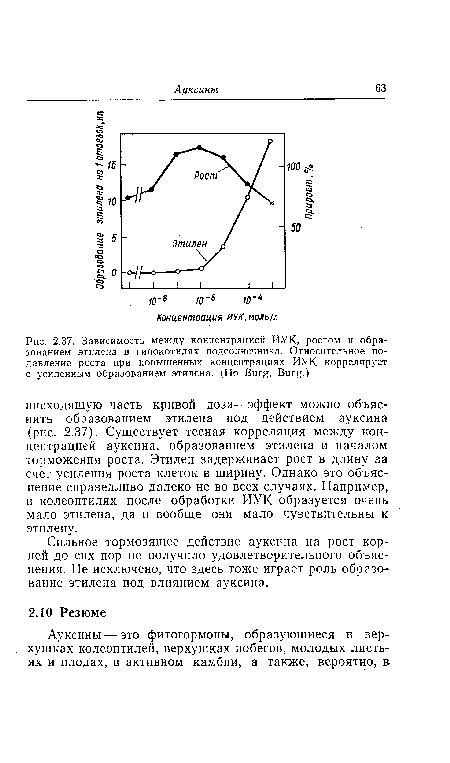
| Зависимость между концентрацией ИУК, ростом и образованием этилена в гипокотилях подсолнечника. Относительное подавление роста при повышенных концентрациях ИУК коррелирует с усиленным образованием этилена. (По Burg, Burg.) |
 |
Далее
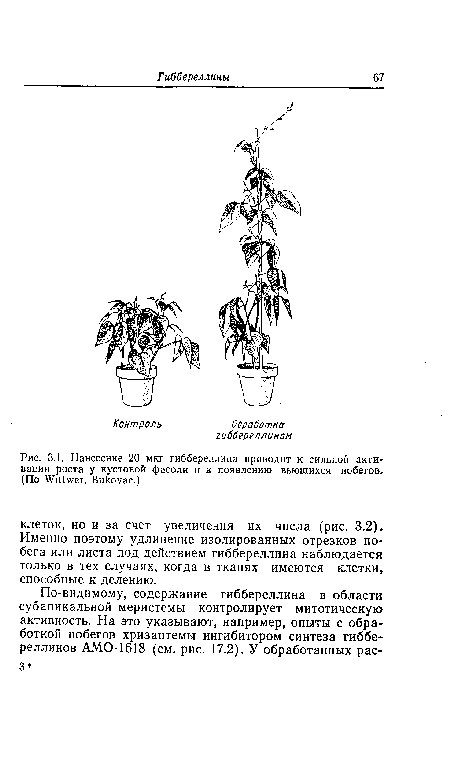
| Нанесение 20 мкг гиббереллина приводит к сильной активации роста у кустовой фасоли и к появлению вьющихся побегов. (По Vittwer, Викоуас.) |
 |
Далее
| Обработка главного побега Бато1и$ гиббереллином в дозе 25 мкг приводит как к усилению роста в длину, так и к увеличению числа клеточных делений. Каждый митоз представлен одной точкой. Показаны продольные срезы главного побега через различные интервалы времени. (По БасИв.) |
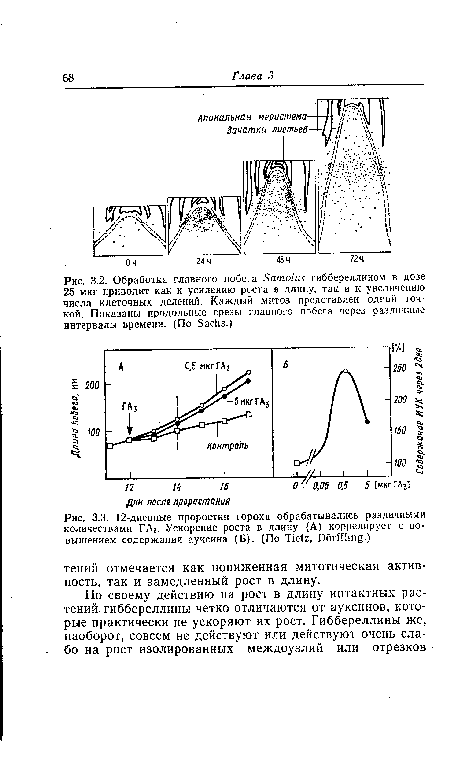 |
Далее
| Схема действия гиб-береллина при прорастании зерна ячменя. Зародыш вырабатывает гиббереллин, который диффундирует к алейроновому слою (а) и здесь индуцирует синтез а-амилазы (б). Новообразованная амилаза гидролизует крахмал эндосперма. Продукты гидролиза крахмала всасываются щитком и поступают в зародыш (в). |
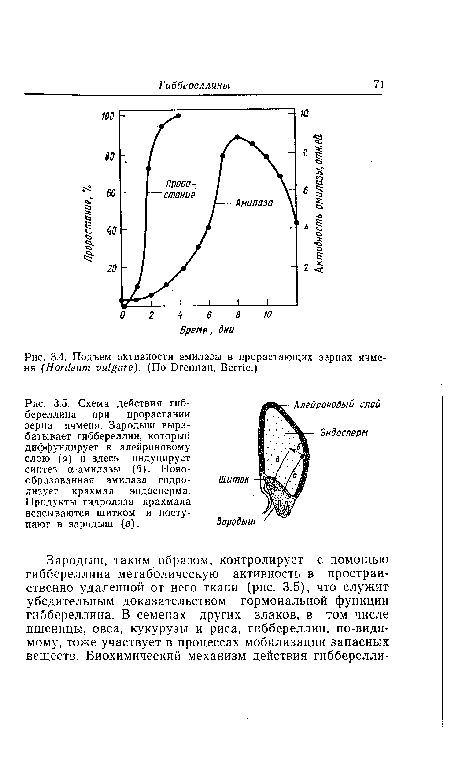 |
Далее
| Известные к 1978 году 52 гиббереллина. (По НесЫеп.) |
 |
Далее
| Важнейшие этапы биосинтеза гиббереллинов. |
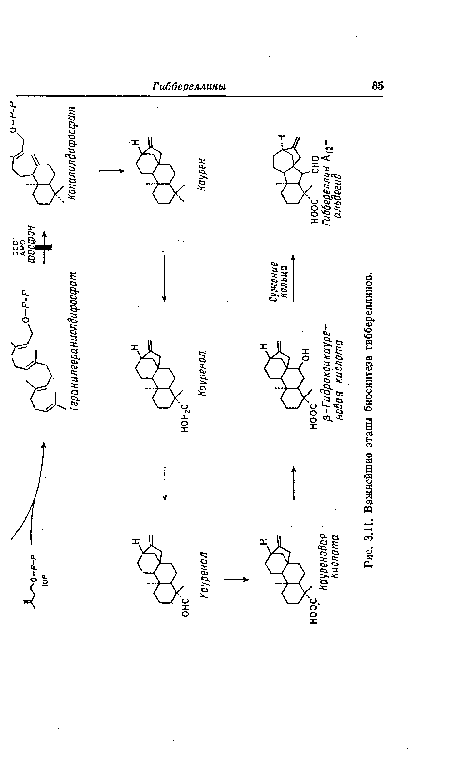 |
Далее
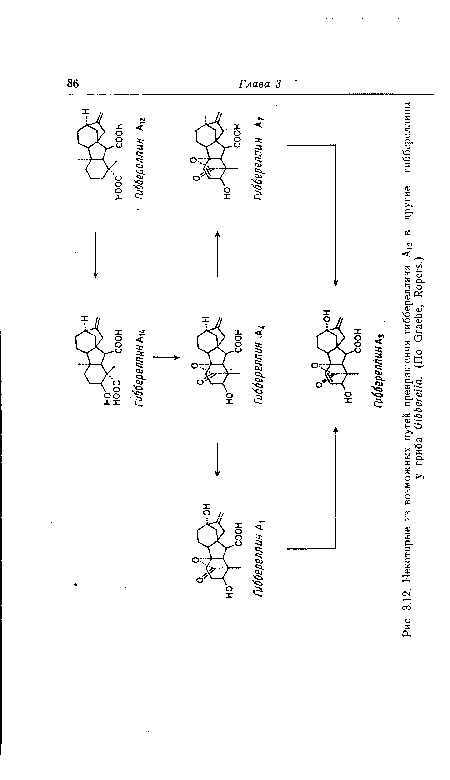
| Некоторые из возможных путей превращения гиббереллина AJ2 в другие гиббереллины |
 |
Далее
| Гиббереллиновая активность в экстрактах из 40 г листьев пшеницы. А. Экстракты из неосвещенных, выросших в темноте листьев. Б. Экстракты из листьев после освещения в течение 20 мин красным светом. Экстракты фракционировали методом тонкослойной хроматографии; активность фракций измеряли в биотесте с проростками салата. После освещения появляются две фракции гиббереллинов. Штриховкой указано значительное ускорение роста. (По Cooke, Saunders.) |
 |
Далее
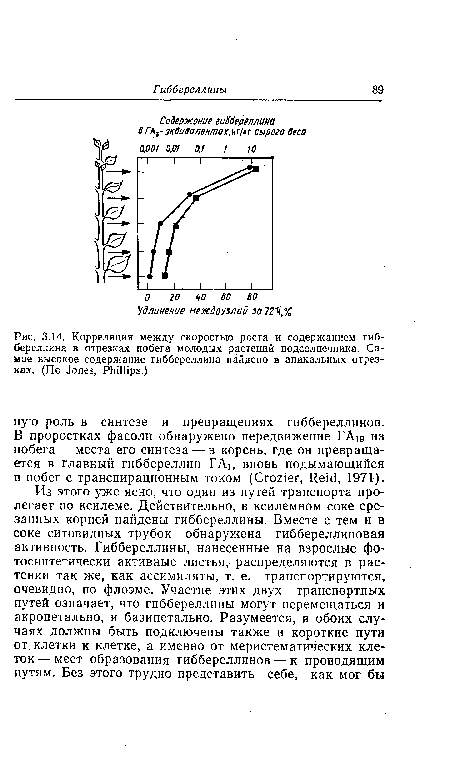
| Корреляция между скоростью роста и содержанием гиб-береллина в отрезках побега молодых растений подсолнечника. Самое высокое содержание гиббереллина найдено в апикальных отрезках. (По Jones, Phillips.) |
 |
Далее
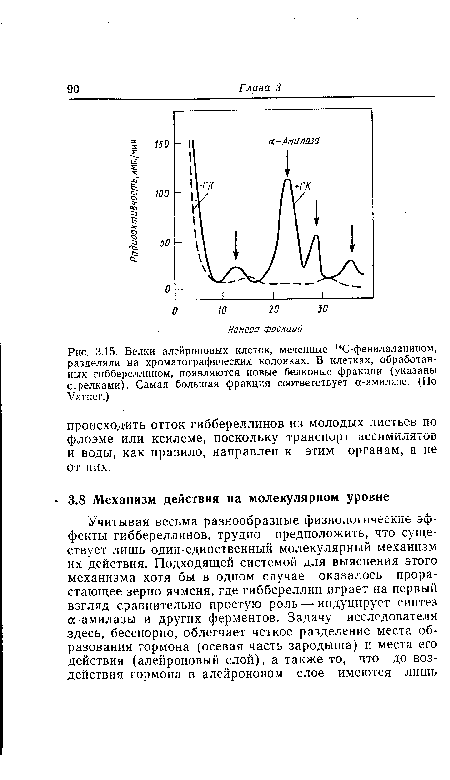
| Белки алейроновых клеток, меченные 14С-фенилаланином, разделяли на хроматографических колонках. В клетках, обработанных гиббереллином, появляются новые белковые фракции (указаны стрелками). Самая большая фракция соответствует а-амилазе. (По Varner.) |
 |
Далее
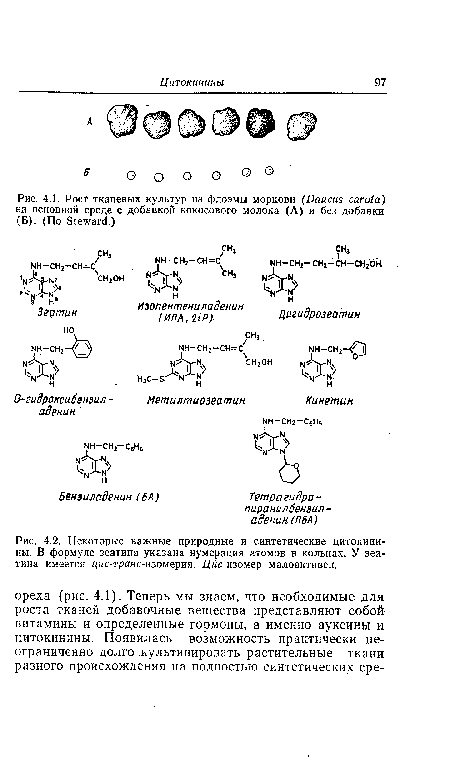
| Рост тканевых культур из флоэмы моркови (Daucus carota) на основной среде с добавкой кокосового молока (А) и без добавки (Б). (По Steward.) |
 |
Далее
| Для роста нормальной ткани Vinca rosea кроме минеральных и органических питательных веществ, а также витаминов необходимы ауксин и цитокинин. Такая зависимость от гормонов слаб» выражена или совсем почти не выражена у опухолевых тканей. Верхний ряд |
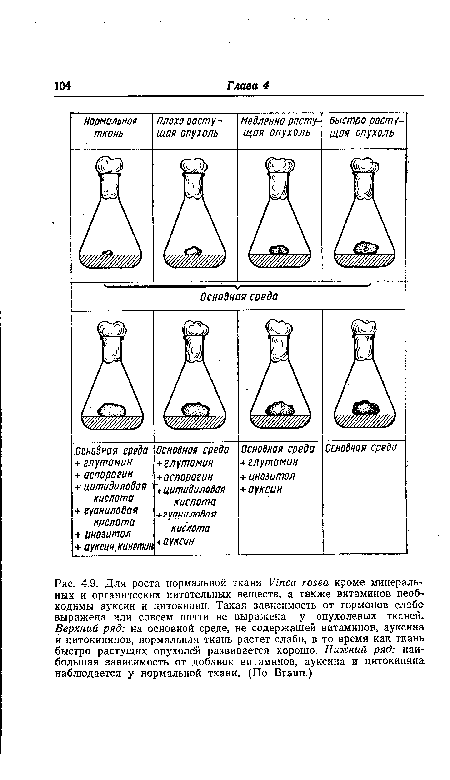 |
Далее
| Табачное растение после многосуточного пребывания в темноте. У нижних листьев (справа) четко выражены признаки старения. Исключение составляет лист (слева), который был опрыскан раствором кине-тина. (По МоШев.) |
 |
Далее
| В экстрактах из плодов двух разных сортов хлопчатника незадолго до преждевременного опадения плодов н во время опадения выявляется повышенная активность ингибиторов (в тесте с отрезками колеоптилей овса). Эти же ингибиторы способствуют опадению листьев. (По АсШсои.) |
 |
Далее
| Структурные формулы абсцизовой кислоты (слева) и ксантоксина (справа). |
|
Далее
| Водный дефицит приводит к значительному повышению уровня АБК в проростках гороха. Содержание воды в нормально обеспеченных водой проростках составляет около 90%; при недостатке воды оно снижается, а содержание АБК увеличивается. |
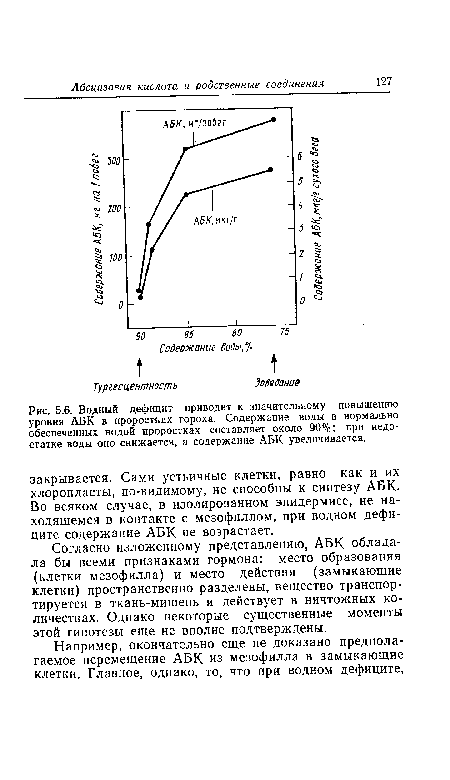 |
Далее
| А. Газовая хроматограмма растительного экстракта. Внесенная проба (1 мкл) содержала около 2-10 |
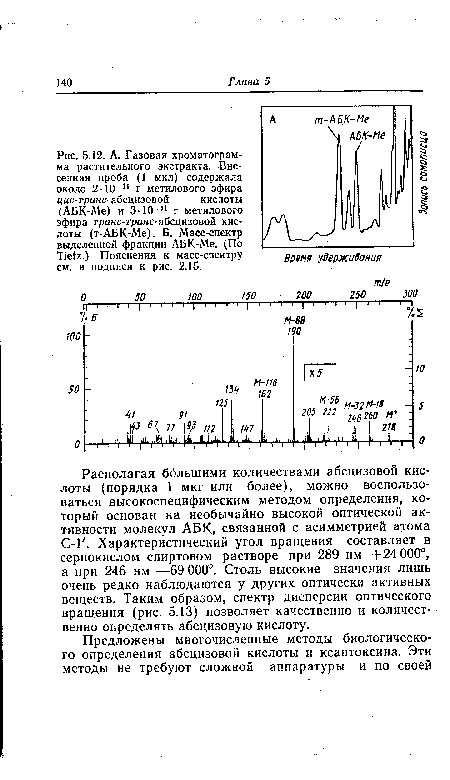 |
Далее
| Спектр дисперсии оптического вращения абсдизовой кислоты (5 мкг/мл), выделенной из плодов томата. Горизонтальная линия относится к растворителю (этанол, подкисленный серной кислотой) . |
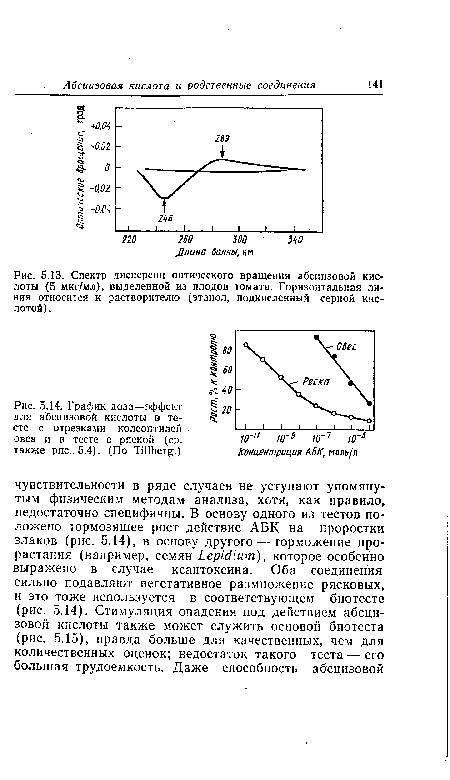 |
Далее
| График доза—эффект для абсцизовой кислоты в тесте с отрезками колеоптилей овса и в тесте с ряской (ср. также рис.. 5.4). (По ТШЬе^.) |
|
Далее
| Некоторые «ингибиторы», регуляторная функция которых, однако, еще сомнительна. |
 |
Далее
| Образование этилена (в мкл на 1 г сырого веса в 1 ч) в различных частях этиолированного, т. е. выращенного в темноте, проростка гороха. (По Burg, Burg.) |
 |
Далее
| Образование этилена во время развития плода дыни. Оно резко увеличивается незадолго до «климактерического» подъема дыхания, указывающего на начало созревания. (По Goesch!, Pratt.) |
 |
Далее
| Если выращенный в темноте проросток гороха в течение минуты освещать красным светом, то апикальная петля временно раскрывается, а образование этилена замедляется. (По ОоеБсЫ.) |
|
Далее
| А. Проростки гороха, выращенные в темноте. Б. Если такие проростки в течение 5 ч освещать красным светом, то апикальная петля распрямляется. В. Опрыскивание этрелом (0,05%; из этрела выделяется этилен) препятствует этому ростовому движению. |
 |
Далее
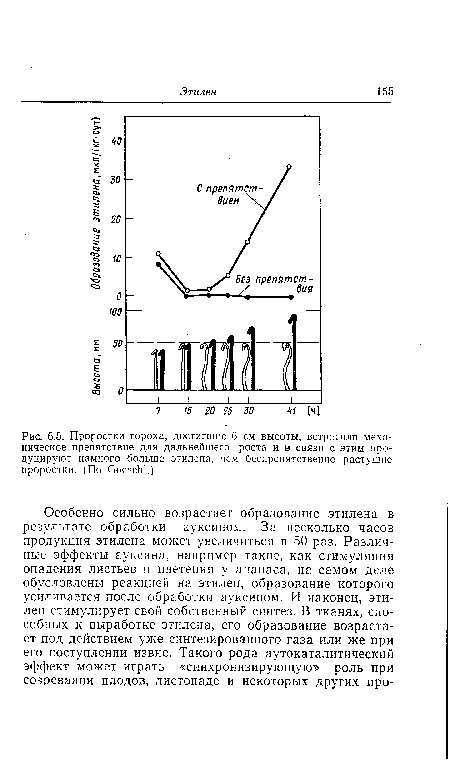
| Проростки гороха, достигшие 6 см высоты, встретили механическое препятствие для дальнейшего роста и в связи с этим продуцируют намного больше этилена, чем беспрепятственно растущие проростки. (По ОоезсЫ.) |
 |
Далее
| А. Влияние этилена на развитие проростков подсолнечника. Проросток развивался из семени, набухавшего в 0,01%-ном растворе этрела (из этрела выделяется этилен). Б. Контрольный проросток. |
 |
Далее
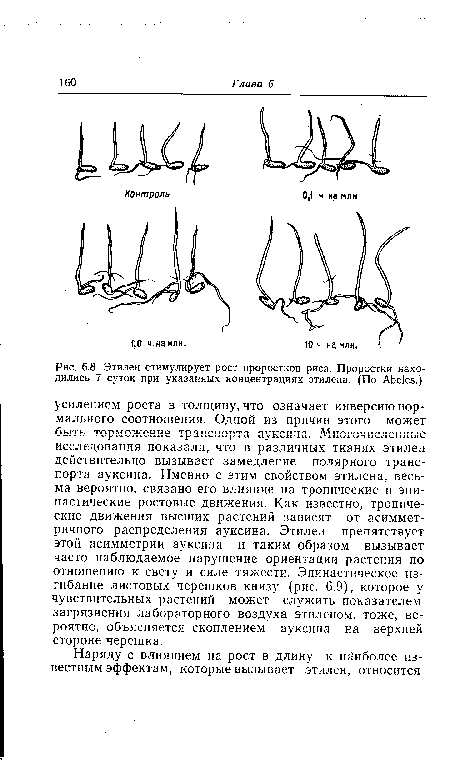
| Этилен стимулирует рост проростков риса. Проростки находились 7 суток при указанных концентрациях этилена. (По Abeles.) |
 |
Далее
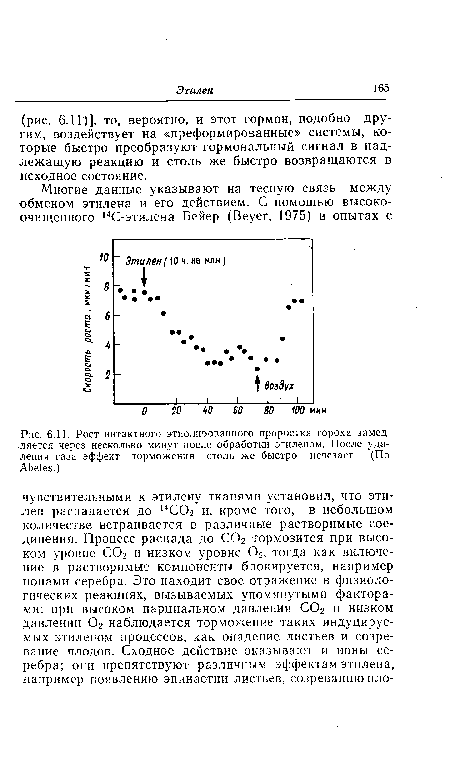
| Рост иитактного этиолированного проростка гороха замедляется через несколько минут после обработки этиленом. После удаления газа эффект торможения столь же быстро исчезает. (По Abeles.) |
 |
Далее
| Эксперимент Чайлахяна с Chrysanthemum, показывающий, что растения способны к образованию цветков лишь в том случае, если листья находились при коротком дне (Б и Г, заштрихованные участки). Интерпретация Чайлахяна |
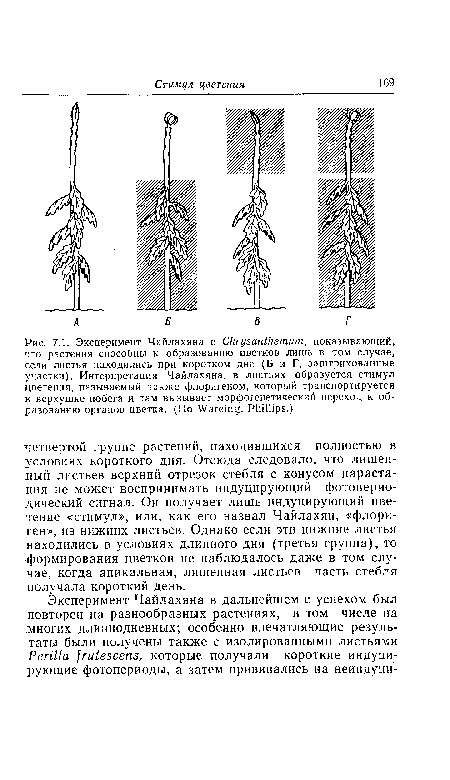 |
Далее
| Растения Xanthium pennsylvanicum с двойным формованным побегом. А. Только один побег (заштрихован) подвергается воздействию короткого индуктивного фотопериода. Б. Для индукции цветения достаточно восьмой доли одного листа. Цветки образуются и на том побеге, который не получал короткого дня. В. Без листьев индукции цветения не происходит. (По Hammer.) |
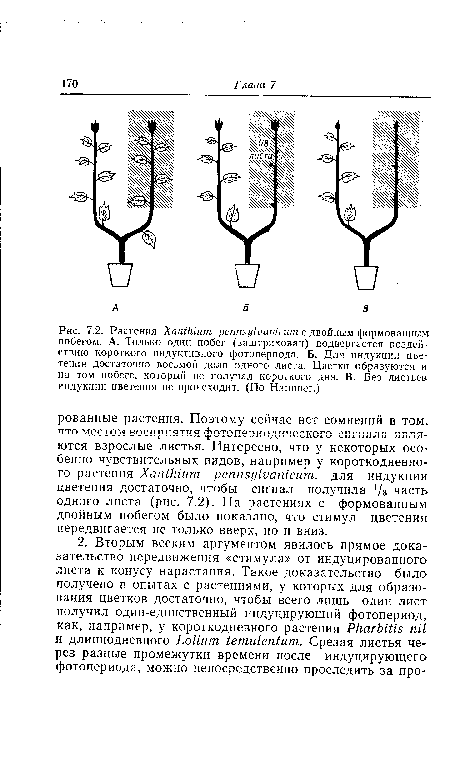 |
Далее
| Переход стимула цветения через место прививки. А. Два растения ХапМит объединены своего рода прививкой; одно из них получает индуктивные короткие фотопериоды, другое остается на длинном дне, но тоже образует цветки. Б. Оба контрольных растения на длинном дне остаются в вегетативном состоянии. |
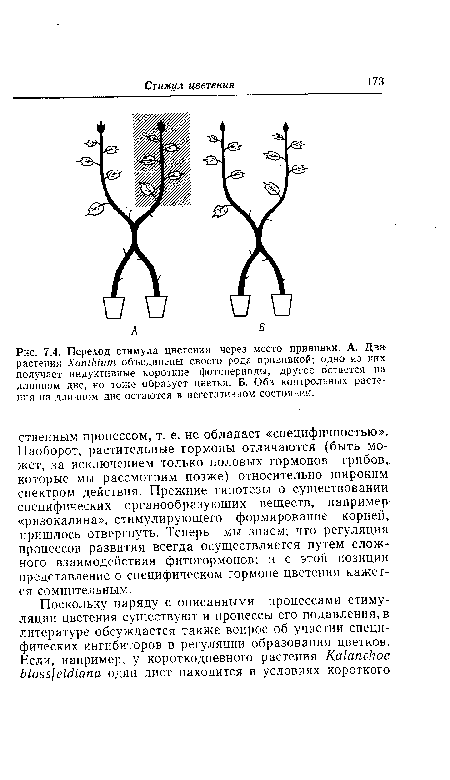 |
Далее
| Триспоровые кислоты В и С — половые гормоны му-коровых грибов. |
 |
Далее
| Гормональные взаимодействия, приводящие к повышению (сплошные стрелки) или понижению (пунктирные стрелки) уровня того или иного гормона. (По ЬеШаш.) |
 |
Далее
| Влияние света на морфогенез проростка гороха. Проросток А выращен в полной темноте, проросток Б — при нормальном освещении. Свет сильно тормозит рост междоузлий, но стимулирует рост листьев. Свет регулирует и многочисленные другие процессы, здесь не показанные, например позеленение и образование механических тканей (выращенные в темноте растения бесцветны и обладают меньшим запасом механической прочности). Для нормального фотоморфогенеза решающее значение имеет спектральный состав света. (По СгШак.) |
 |
Далее
| Развитие плодов яблони. А — нормальное развитие; Б — после обработки гибберелловой кислотой (5■10_3 моль/л) без предварительного опыления; В — после обработки экстрактом семян. (По Dennis.) |
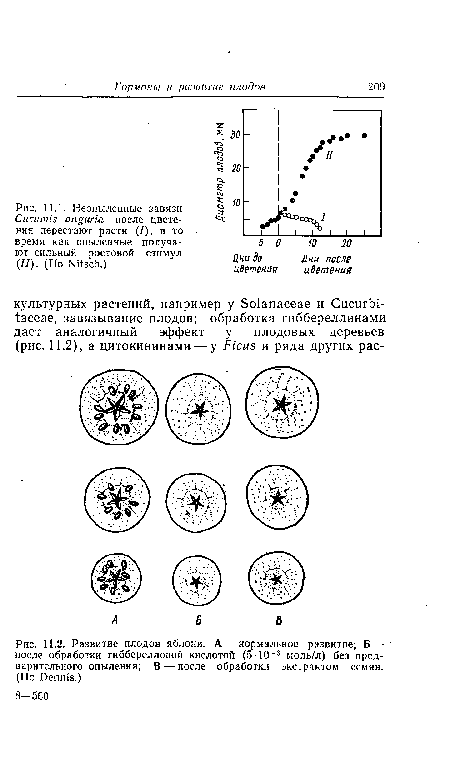 |
Далее
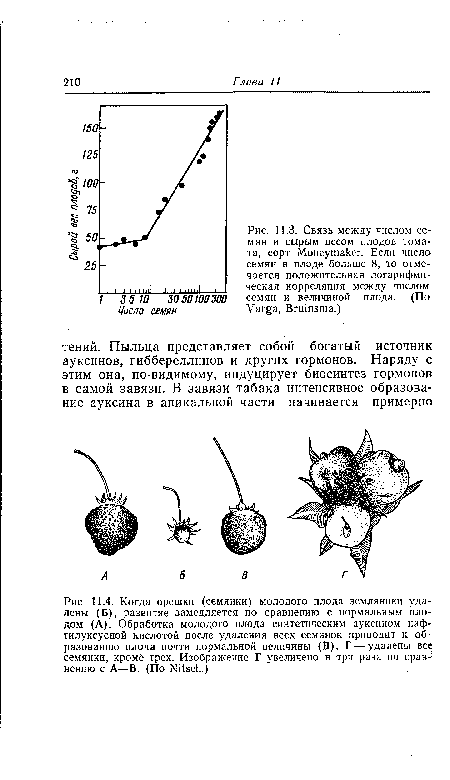
| Связь между числом семян и сырым весом плодов томата, сорт Moneymaker. Если число семян в плоде больше 8, то отмечается положительная логарифмическая корреляция между числом семян и величиной плода. (По Varga, Bruinsma.) |
 |
Далее
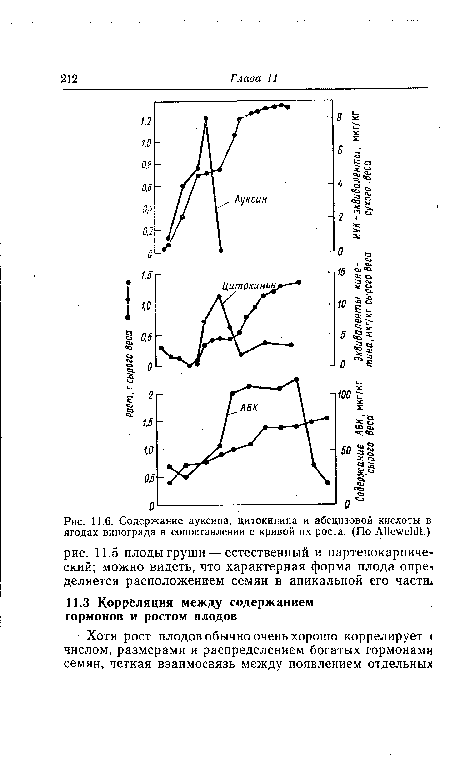
| Когда орешки (семянки) молодого плода земляники удалены (Б), развитие замедляется по сравнению с нормальным плодом (А). Обработка молодого плода синтетическим ауксином наф-тилуксусной кислотой после удаления всех семянок приводит к образованию плода почти нормальной величины (В). Г — удалены все семянки, кроме трех. Изображение Г увеличено в три раза по срав- |
|
Далее
| Влияние семян на форму плода. Л — плод с семенами. Б — бессемянный плод груши сорта Konferenz; он более удлиненный по сравнению с природными, формами, содержащими семена. (По Luckwill.) |
 |
Далее
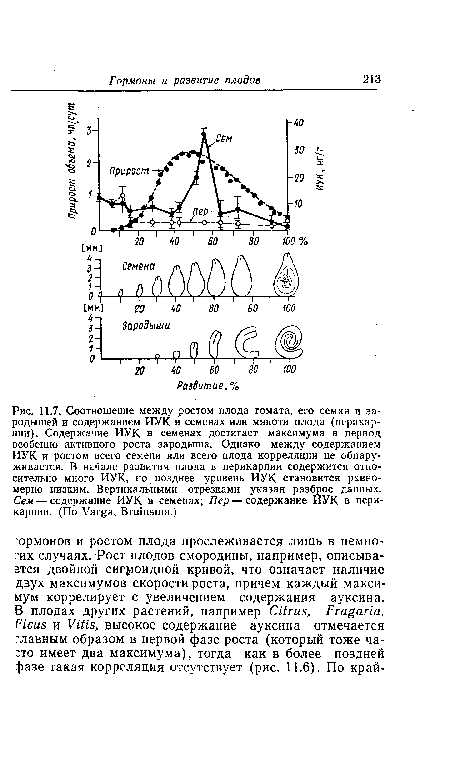
| Содержание ауксина, цитокинина и абсцизовой кислоты в ягодах винограда в сопоставлении с кривой их роста. (По А11е>теШ.) |
 |
Далее
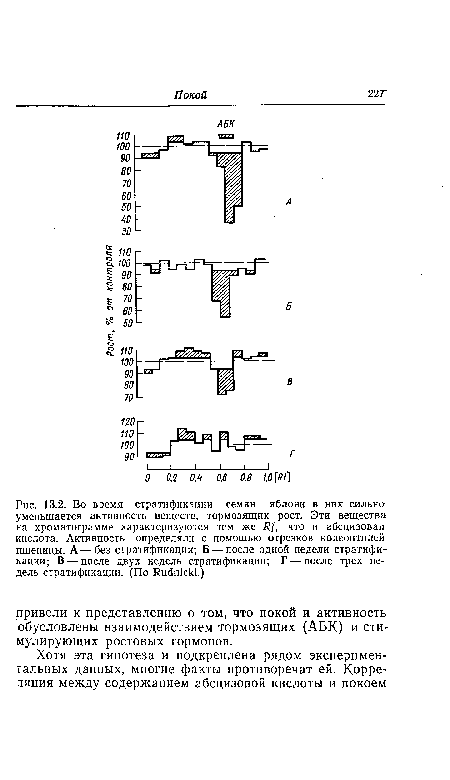
| Соотношение между ростом плода томата, его семян и зародышей и содержанием ИУК. в семенах или мякоти плода (перикарпии). Содержание ИУК в семенах достигает максимума в период особенно активного роста зародыша. Однако между содержанием ИУК и ростом всего семени или всего плода корреляции не обнаруживается. В начале развития плода в перикарпии содержится относительно много ИУК, но позднее уровень ИУК становится равномерно низким. Вертикальными отрезками указан разброс данных. Сем — содержание ИУК в семенах; Пер — содержание ИУК в перикарпии. (По Varga, Bruinsma.) |
 |
Далее
| Схематическое изображение отделительной ткани листового черешка, (по АсШсой.) |
 |
Далее
| Временной ход отделения листовых черешков Coleus после удаления листовых пластинок. Если верхушка побега сохранена, то за 10 дней отделяется 100% листовых черешков (/). Если верхушка побега удалена, опадение листьев замедляется (II). Нанесение ИУК-пасты на поверхность среза компенсирует отсутствие верхушки побега (III). (По 10 20 30 [Пни] Jacobs.) |
![Временной ход отделения листовых черешков Coleus после удаления листовых пластинок. Если верхушка побега сохранена, то за 10 дней отделяется 100% листовых черешков (/). Если верхушка побега удалена, опадение листьев замедляется (II). Нанесение ИУК-пасты на поверхность среза компенсирует отсутствие верхушки побега (III). (По 10 20 30 [Пни] Jacobs.)](/static/pngsmall/798445344.png) |
Далее
| Во время стратификации семян яблони в них сильно уменьшается активность веществ, тормозящих рост. Эти вещества на хроматограмме характеризуются тем же что и абсцизовая кислота. Активность определяли с помощью отрезков колеоптилей пшеницы. А — без стратификации; Б — после одной недели стратификации; В — после двух недель стратификации; Г — после трех недель стратификации. (По Кис!шск1.) |
 |
Далее
| Активность ауксина и цитокинина в соке ксилемы у сеянцев яблони минимальна в период покоя почек и максимальна в период, предшествующий росту, или во время роста. Жирная кривая —• эквиваленты ИУК; тонкая кривая — эквиваленты кинетина. (По Luckwill, White.) |
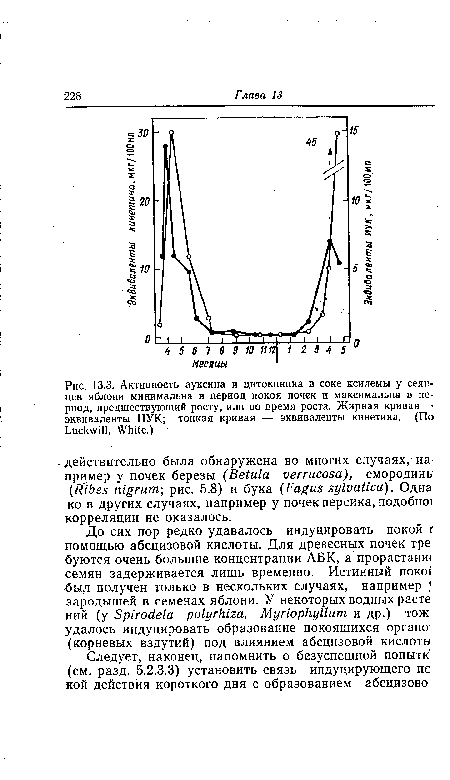 |
Далее
| Пример корреляций |
 |
Далее
| Пример коррелятивных взаимоотношений. А. Столоны дикого картофеля, Solanum andigena, растут горизонтально. Б. После декапитации главного побега столоны выпрямляются и вместо чешуйчатых листьев образуют нормальные. Диагеотропизм, таким образом, контролируется верхушечной почкой главного побега. (По Booth.) |
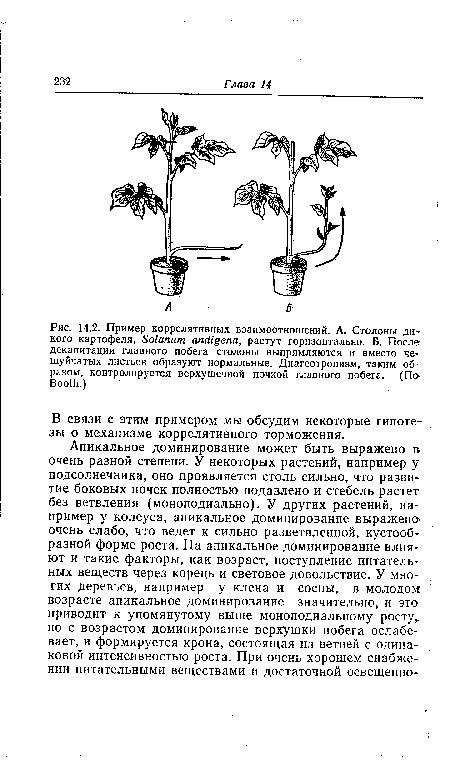 |
Далее
| Как у побегов, так и у корней геотропический изгиб начинается у верхушки органа и распространяется базипетально, причем верхушка опять распрямляется. У побегов нередко сначала возникают перегибы, которые выправляются за счет встречной реакции. Отметки на верхней стороне корня показывают, что главная зона роста находится около его кончика. |
 |
Далее
| Если у вертикально ориентированного корня кукурузы, сорт КеЬ^оп, удалить корневой чехлик, то рост ускоряется по сравнению с интактным корнем, по крайней мере в первые 3 часа. (По РПе1.) |
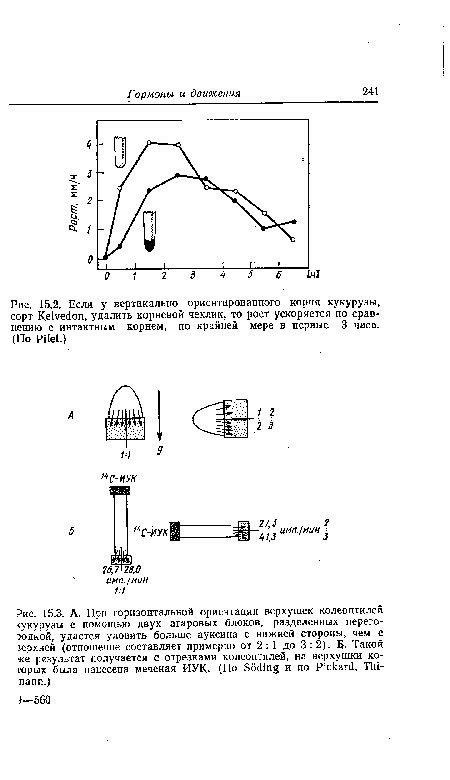 |
Далее
| А. При горизонтальной ориентации верхушек колеоптилей кукурузы с помощью двух агаровых блоков, разделенных перегородкой, удается уловить больше ауксина с нижней стороны, чем с зерхней (отношение составляет примерно от 2 |
|
Далее
| А. Проросток подсолнечника из горизонтального положения геотропически изгибается кверху. Б. Такой же проросток, обработанный ингибитором транспорта ауксина — морфактином, не проявляет геотропической реакции. (По Ворр.) |
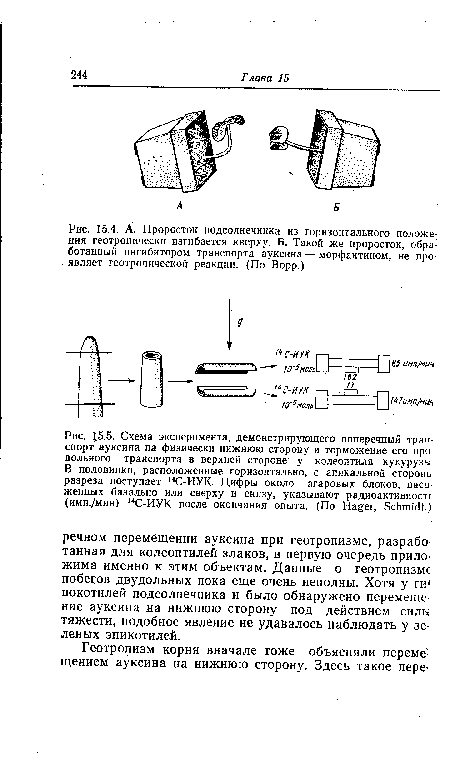 |
Далее
| Фототропический изгиб колеоптиля овса после освещения (4 с, 30 лк). Изгиб начинается от верхушки и распространяется бази-петально, причем верхушка снова распрямляется. (По Апэг.) |
 |
Далее
| Участок графика доза — эффект для фототропического изгиба проростков чечевицы и колеоптилей овса. Максимумы первого положительного изгиба у обоих видов совпадают. Отрицательный изгиб при 104 лк-с наблюдается только у овса. Второй и третий максимумы положительных изгибов не представлены. (По 81еуег, ЫЬЬей.) |
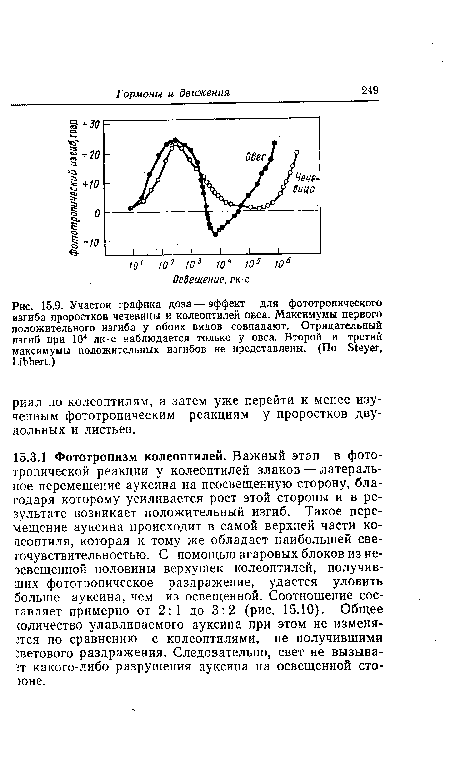 |
Далее
| Доказательство поперечного передвижения ауксина в верхушках колеоптилей кукурузы после фототропического раздражения. Верхушки колеоптилей, выращенных в темноте, помещали н? агаровые блоки и затем подвергали одностороннему освещению (В-И). После этого определяли содержание ауксина в блоках (тест на изгиб колеоптилей овса). Цифры под блоками — изгиб а градусах. Вертикальными линиями, проведенными через колеоптили или блоки, показаны стеклянные перегородки. А—Г. Содержание ауксина в агаровых блоках практически одинаково. Количестве отдаваемого ауксина не изменяется ни после освещения, ни после введения стеклянного барьера. Д, Е. Если перегородка делит толькс агаровый блок, то в неосвещенной половине колеоптиля обнаружи- |
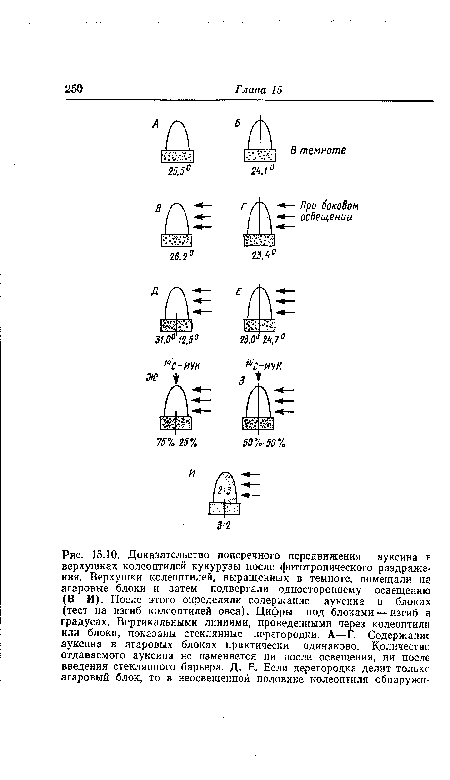 |
Далее
| Фототропическая память». В проростках подсолнечника после декапитации содержание ауксина падает. Затем их освещают с одной стороны. Однако фототропический изгиб происходит только тогда, когда на гипокотиль наносят ауксин, причем это возможно еще через 3 ч после выключения света. (По Brauner, Rau.) |
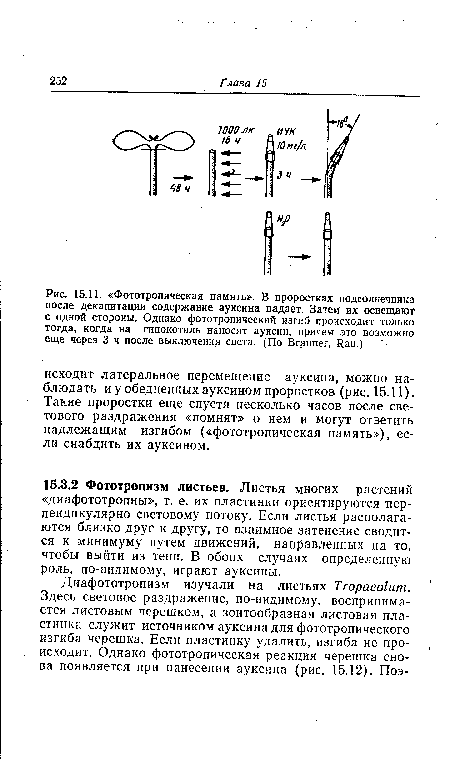 |
Далее
| Если одну половину листа Лг^га затенить, то лист изгибается так, чтобы выйти из тени. Из затененной половины в агаровый блок переходит больше ауксина, чем из незатененной. (По Уашапе.) |
 |
Далее
| Влияние семядолей на реакцию изгиба гипокотилей подсолнечника. А, Б. Если одну из семядолей затенить, гипокотиль изгибается в сторону освещенной семядоли. В. Из затененного гипо-котиля в агаровый блок переходит больше ауксина, чем из освещенного. (По Lam, Leopold.) (Этот результат можно объяснить и по-иному |
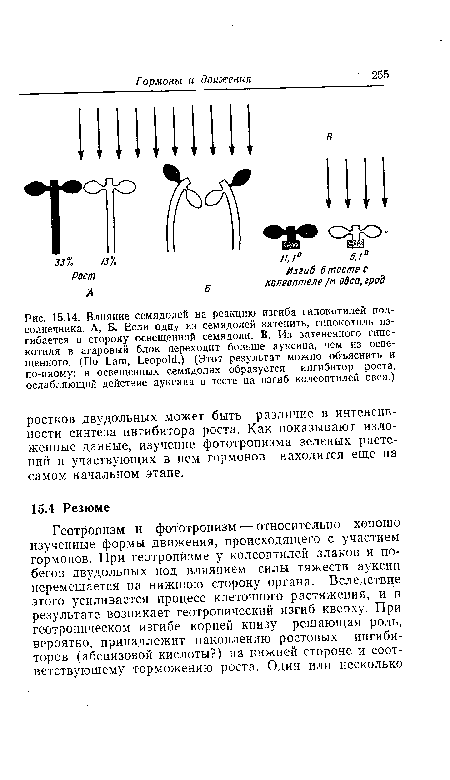 |
Далее
| Некоторые регуляторы роста, имеющие практическое значение. |
 |
Далее
| Структурные формулы двух регуляторов роста, практическое значение которых не столь велико, — фосфона Д (слева) и АМО-1618 (справа). |
|
Далее
| А. Нормальные проростки гороха (контроль). Б. Проростки гороха с нарушенным морфогенезом после 12-часовой обработки семян флуоренолом (морфактином) в концентрации 5-10-5 моль/л. Рост побега замедляется, боковые почки трогаются в рост. Наряду с этим удлиняется главный корень и подавляется образование боковых корней. (По Ziegler.) |
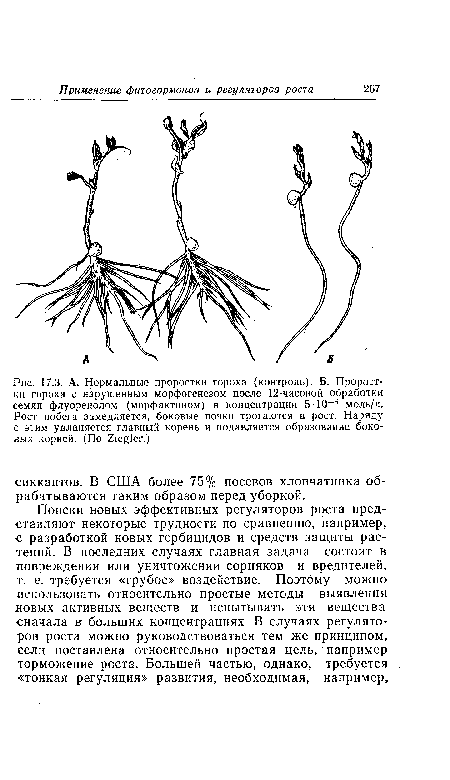 |
Далее
| А. Черенки айвы (контроль). Б. Черенки айвы через 3 недели после 24-часовой обработки индолил-3-масляной кислотой в концентрации 20 мг/л. (По Реагзе.) |
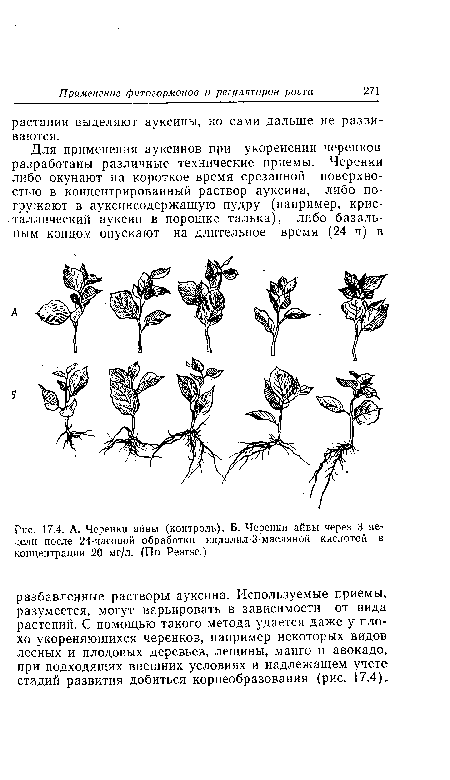 |
Далее
| Селективное действие гербицида, связанное с различным расположением конусов нарастания. А — защищенное положение; Б — незащищенное положение. (По Weaver.) |
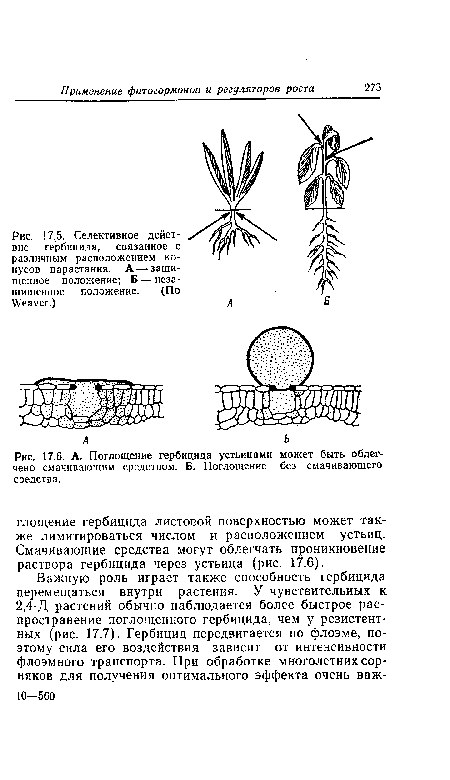 |
Далее
| А. Поглощение гербицида устьицами может быть облегчено смачивающим средством. Б. Поглощение без смачивающего средства. |
|
Далее
| Селективное действие гербицида, связанное с легкостью его передвижения в растении. А. 2,4-Д медленно распространяется у злаков. Б. Напротив, у широколистных двудольных сорняков 2,4-Д распространяется быстро. Стрелки указывают место нанесения гербицида, точки —-• его распределение. (По Weaver.) |
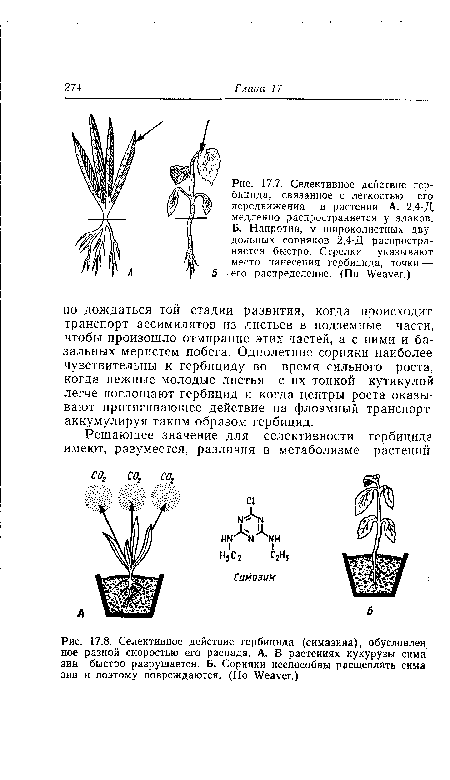 |
Далее
| Селективное действие слабого гербицида (2,4-ДМ), связанное с его превращением в высокотоксичное соединение (2,4-Д). А. Устойчивое культурное растение де способно к р-окислению |
 |
Далее
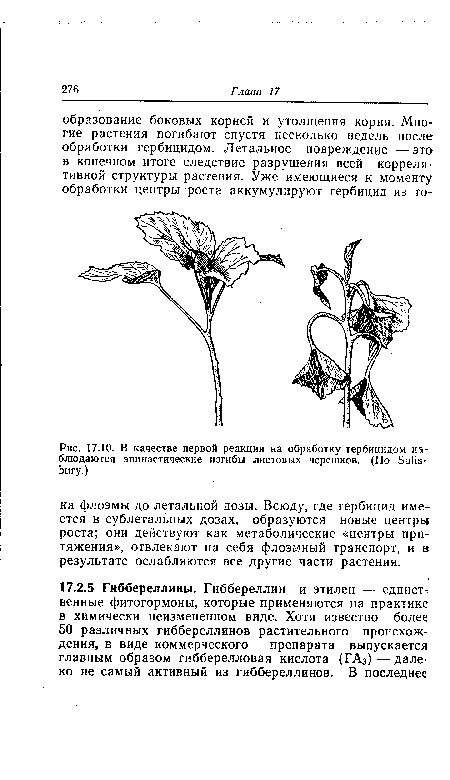
| В качестве первой реакции на обработку гербицидом наблюдаются эпинастические изгибы листовых черешков. (По Salisbury.) |
 |
Далее