ЦТК) состоит из серии последовательных реакций, катализируемых десятью различными ферментами. Последовательность реакции описана ниже.[ ...]
Цикл Кребса подвержен регуляции; его скорость зависит от потребности в АТФ других метаболических реакций.[ ...]
Кребса цикл — серия ферментативных реакций, конвертирующих пиро-виноградную кислоту в двуокись углерода, воду и энергию.[ ...]
| Цикл трикарбоновых кислот (цикл Кребса). | 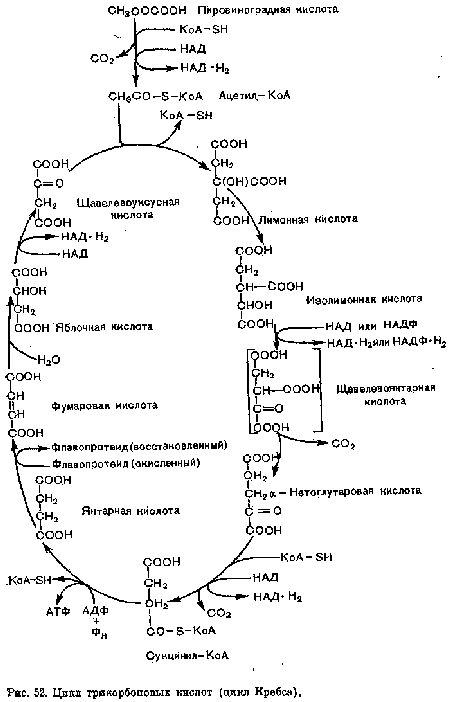 |
| Н,0 Цикл Кребса -- — |  |
| Схема цикла трикарбоновых кислот или цикл Кребса | 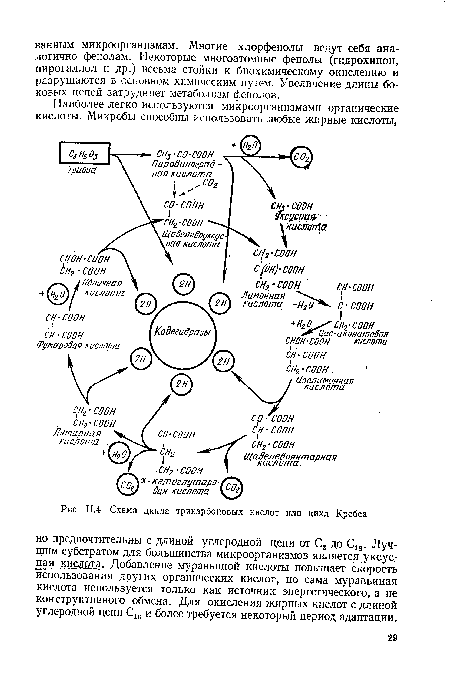 |
В 1935 г. венгерский ученый А. Сент Дьердьи установил, что добавление небольших количеств органических кислот (фумаровой, яблочной или янтарной) усиливает поглощение кислорода измельченными тканями. Продолжая эти исследования, Г. Кребс пришел к выводу, что главным путем окисления углеводов являются циклические реакции, в которых происходит постепенное преобразование ряда органических кислот. Эти преобразования и были названы циклом трикарбоновых кислот или циклом Кребса.[ ...]
Биологическое значение цикла Кребса заключается не только в том, что он является завершающим этапом в генерировании энергии, но и в том, что он «поставляет промежуточные продукты для биосинтеза.[ ...]
Янтарная кислота, образовавшаяся в цикле Кребса, дает основу для.образования порфиринового ядра хлорофилла. Поскольку имеется ряд реакций и процессов, благодаря которым отдельные компоненты извлекаются из цикла Кребса, должны быть и обратные процессы, поставляющие их в цикл. Если бы этого не было, скорость превращения в аэробной фазе дыхания заметно бы снизилась. Такими реакциями является окислительное дезаминирование аминокислот, приводящее к образованию органических кислот.[ ...]
Последняя стадия заключается в преобразовании кислот в цикле Кребса до простых продуктов, при полном окислении до диоксида углерода и воды. Наиболее энергично окисляют алканы бактерии Pseudomonas, микобактерии, актиномицеты. Разложение углеводородов может происходить и в анаэробных условиях в присутствии веществ, способных отдавать кислород.[ ...]
Не менее важное значение имеют и промежуточные продукты цикла Кребса. Благодаря реакции переампнирования эти кислоты могут быть источником аминогруппы для других аминокислот и таким образом- являться важнейшими промежуточными продуктами для синтеза как белка, так и пуриновых и пиримидиновых азотистых оснований.[ ...]
В результате четырех окислительно-восстановительных реакций цикла Кребса происходит перенос трех пар электронов над НДЦ и одной пары электронов на ФАД. Восстановленные этим путем переносчики электронов НАД и ФАД подвергаются затем окислению уже в цепи переноса электронов, в результате которого генерируется одиннадцать молекул АТФ. Одна высокоэнергетическая связь генерируется непосредственно в цикле Кребса. Таким образом, на каждый двухуглеродный фрагмент, полностью окисляемый до Н20 и С02, генерируется двенадцать высокоэнергетических фосфатных связей.[ ...]
Таким образом, происходит, в частности, полное окисление глюкозы до С02(в цикле Кребса) и Н20 (в цепи переноса электронов). В результате за счет окисления одной молекулы глюкозы происходит образование 38 молекул АТФ, механизм действия которых как энергетического челнока описан ранее. В клетках окислению подвергаются также другие сахара, жиры и аминокислоты.[ ...]
Интенсивность дыхания — среднее из суммы многочисленных реакций гликолиза и цикла трикарбоновых кислот (цикла Кребса). Поэтому для нее верно среднее значение (¿ю от 2 до 3 в биологически важном диапазоне температур от 0 до 30 °С.[ ...]
Окисление жирных кислот идет через стадии образования других кислот, которые вступают в цикл Кребса. Органические кислоты могут использоваться микроорганизмами в качестве источника энергии и как материал для биосинтеза. Исключение составляет муравьиная кислота, которая участвует только в катаболитическом процессе.[ ...]
Продуцируемая при р-окислении уксусная кислота окисляется до воды и диоксида углерода в известном цикле три- и дикарбоновых кислот (в цикле Кребса).[ ...]
Третья стадия — электронно-транспортная цепь (ЭТЦ). В процессе окисления пировипоградыой кислоты в цикле Кребса образовались пары водорода 2Н, которые мы можем рассматривать как 2Н++2ё. Именно в таком виде опи и передаются по цепи перепосчиков. Движущей силой транспорта водорода в дыхательной цепи является разность потенциалов. Это и позволяет электронам передвигаться по направлению к кислороду (паивыс-ший положительный потенциал). Оксидоредуктазы делятся па три главных подкласса.[ ...]
Марганец поступает в растение в виде ионов Мп34. В связи с этим понятно болы ое значение марганпа для процесса дыхания, особенно его аэробной фазы. Правда, имеются данные, что в некоторых из етих реакций марганец может быть заменен кобальтом.[ ...]
Таким образом, основное количество энергии запасается именно на стадии окисления в ЦТК. Замечательно то, что цикл Кребса универсален. Такой тип окисления характерен и для простейших, и для бактерий, и для клеток высших животных и растений.[ ...]
Суммарно при полном окислении моля глюкозы образуется 38 молекул АТФ. Из них 24 при окислении двух молекул ПВК в цикле Кребса.[ ...]
Относительная роль этих путей дыхапия может меняться в зависимости от типа растений, возраста, фазы развития, а также в зависимости от условии внешней среды. Процесс дыхания растений осуществляется во всех ваешпих условиях, в которых возможна жизнь. Растительный организм не имеет приспособлений к регуляции температуры, поэтому процесс дыхания осуществляется при температуре от —50 до +50вС. Нет приспособлений у растений и к поддержанию равномерного распределения кислорода по всем тканям. Именно необходимость осуществления процесса дыхания в разнообразных условиях привела к выработке в процессе эволюции разнообразных путей дыхательного обмена и к еще большему разнообразию ферментных систем, осуществляющих отдельные этапы дыхания. При этом важно отметить взаимосвязь всех процессов обмена в растительном организме. Изменение пути дыхательного обмена приводит к глубоким изменениям во всем метаболизме растительных организмов.[ ...]
Важное значение имеет регулирование с помощью конечных продуктов реакций. Это происходит при замедлении подачи активного ацетата и предупреждает полное превращение всех интермедиатов цикла в ЩУК.[ ...]
Быстрое связывание поступивших в корни ионов аммония возможно при одновременном притоке углеводов из листьев. Сахара благодаря гликолизу и циклу Кребса (превращение десяти органических кислот) переходят в кетокислоты. Последние подвергаются ферментативному аминиро-ванию за счет аммиака и Дают ряд аминокислот.[ ...]
В дальнейших этапах окисления электроны, отщепляемые от богатых энергией промежуточных соединений в окислительновосстановительных реакциях цикла Кребса, поступают в цепь переноса электронов и передаются по ней, попадая на каждом новом этапе переноса на все более низкий энергетический уровень до тех пор, пока в конце концов не будут присоединены кислородом, восстанавливающимся в результате этого до воды. При передаче электронов по цепи от одного переносчика к другому - на все более низкий энергетический уровень - значительная часть их свободной энергии запасается в процессе синтеза АТФ из АДФ и неорганического фосфата... Таким путем создается резерв АТФ, который необходим для работы клетки.[ ...]
У тли показано (3) отсутствие гексекиназной системы, но несмотря на это, гексозофосфаты быстро используются. Это происходит в результате прямого окисления глюкозы по пентозному циклу. Известно, (4), что в случаях, когда цикл Кребса служит не для производства энергии, а главным образом для синтеза аминокислот или других важных метаболитов, пентозный цикл может функционировать в некоторых организмах как большой источник энергии. Вполне вероятно, что тля такой организм, где пентозный цикл является большим источником энергии, а фосфорилирование в этом случае сопряжено с окислением пиридиннуклеотида.[ ...]
В последующей реакции происходит окисление малата, что приводит к регенерированию оксалоацетата (С4).[ ...]
Следовательно, «топливом» для окисления в митохондриях являются пируват и жирные кислоты. Ацетил-КоА обладает высоким потенциалом переноса ацетильных групп. Следовательно, топливные молекулы вступают в цикл Кребса в виде ацетил-КоА. Непрерывность же снабжения окислительных процессов «топливом» обеспечивается запасанием животными клетками липидов, являющихся главным ресурсом жирных кислот, а также гликогена, являющегося источником глюкозы.[ ...]
На протяжении всего цикла агрессивность изменяется, хотя, каким образом, не вполне ясно. По мнению Кребса (Krebs, 1985), она максимальна в фазе роста популяции, в ходе же снижения численности животные малоагрессивны. В то же время Варковска-Дратнал и Стенсет (Warkowska-Dratnal, Stenseth, 1985) утверждают, что агрессивность минимальна в начале роста популяции и максимальна, когда плотность еще высока, но ее максимум уже позади.[ ...]
У некоторых растений, таких, как Dichapetalum spp. в Южной Африке и Acacia georginae Gidyea, в Австралии (Peters, Schorthouse, 1964; Peters et al., 1965), происходит образование фторуксусной кислоты (FCH2COOH), что может приводить к гибели пастбищных животных. Относительно неядовитая фторуксусная кислота под действием ферментов организма животного превращается во фторцитрат, который ингибирует реакции окисления лимонной кислоты в цикле Кребса (Peters et al., 1953). О потенциальной токсичности фторацетата можно судить по величине LD50, равной 1 мг/кг веса тела для морских свинок.[ ...]
На этой точке зрения особенно настаивает Маргулис (Margulis, 1975). Позднее поглощение клеток сине-зеленых водорослей могло бы привести к появлению автотрофного «организма», предка всего царства растений. Эта последовательность событий является чисто гипотетической и вызывает много скептических замечаний, но, по крайней мере отчасти, она может соответствовать действительности. До сих пор существуют близкие к предполагаемым симбионтам формы, включая бактерию Paracoccus denitrificans, рядом признаков напоминающую гипотетического свободножи-вущего предшественника митохондрии. Если теория Маргулис верна, то основная часть этой главы была посвящена второй стадии развития мутуализма, когда все более интегрируются друг с другом пары видов, каждый из которых по происхождению является симбиотической ассоциацией.[ ...]
Унц и Дондеро [502, 503] разработали метод выделения и изолировали из активных илов 65 штаммов Z. ramigera. Авторы детально изучили их физиолого-биохимические свойства и показали, что эти культуры растут в широком диапазоне температур от 9 до 37° С. Оптимальными являются температура 28° и pH 7,0. Бактерии не растут в анаэробных условиях, но хорошо переносят их в течение 24 дней. При исследовании отношения выделенных штаммов к различным источникам углерода установлено, что крахмал, амигдалин, инулин, некоторые первичные спирты, насыщенные жирные кислоты и интермедиаты цикла Кребса поддерживают рост большинства изолятов. Аспарагин, аспарагиновая и глутаминовая кислоты пригодны для большей части культур как единственный источник углерода и азота. Авторы считают, что штаммы рода Zoogloea доминируют в природных хлопьевидных зооглейных колониях. Из 38 штаммов, выделенных из пены, 22 изолята принадлежали к роду Zoogloea.[ ...]
О том, что дефицит кислорода в семени может служить фактором, обусловливающим покой, предположили на основе работ с семенами риса, которые входят в состояние покоя сразу же после их уборки, а в процессе хранения постепенно выходят из этого состояния. Находящиеся в покос семена риса можно индуцировать к прорастанию удалением пленки, что указывает на важное значение оболочки этих семян. Хранение в атмосфере кислорода значительно сокращает период покоя, по-видимому, вследствие включения некоторых окислительных реакций в процессы дозревания во время храпения. Однако Робертс, испытавший действие различных ингибиторов дыхания (в том числе ингибиторов терминального окисления, цикла Кребса и гликолиза), получил неожиданный результат: оказалось, что эти ингибиторы стимулируют прорастание покоящихся семян риса. Робертс предположил, что, прежде чем наступит прорастание, должно произойти несколько окислительных реакций и что эти реакции конкурируют за кислород с процессами дыхания, включающими гликолиз, цикл Кребса и систему терминального окисления в условиях низкого уровня кислорода, содержащегося в семенах; следовательно, при ингибировании этих процессов дыхания различными веществами будет высвобождаться большое количество кислорода для других окислительных реакций, которые, по предположению Робертса, могут включать «пентозофосфатный путь» углеводного метаболизма. Имеются достаточно обоснованные данные, что прекращение покоя у семян некоторых видов сопровождается переключением с гликолитического пути на пентозофосфатный. Далее предполагается, что действие ингибиторов дыхания в стимуляции прорастания осуществляется через ингибирование фермента катал азы, которая расщепляет перекись водорода. Сохраненная таким путем перекись увеличивает, как предполагается, активность пеитозофосфатного пути.[ ...]
Специфика биологических макромолекул определяется также и тем, что процессы биосинтеза осуществляются в результате одних и тех же этапов метаболизма. Больше того, биосинтезы нуклеиновых кислот, аминокислот и белков протекают по сходной схеме у всех организмов независимо от их видовой принадлежности. Универсальными являются также окисление жирных кислот, гликолиз и другие реакции. Например, гликолиз происходит в каждой живой «летке всех организмов-эукариотов и осуществляется в результате 10 последовательных ферментативных реакций, каждая из которых катализируется специфическим ферментом. Все аэробные организмы-эукариоты обладают молекулярными «машинами» в их митохондриях, где осуществляется цикл Кребса и другие реакции, связанные с освобождением энергии. На молекулярном уровне происходят многие мутации. Эти мутации изменяют последовательность азотистых оснований в молекулах ДНК.[ ...]
Биологическое сгорание сахара совершается путем чрезвычайно сложных серий реакций, в которые вступают многие специфические ферментные системы и носители энергии. В последовательных реакциях от шестиуглеродного сахара до углекислоты фосфорилированные сахара превращаются в трехуглеродную пировиноградную кислоту (СН3СОСООН). Эта реакция, известная как гликолиз, свойственна многим организмам. Организмы, которым для существования не требуется кислород (анаэробы), превращают пировиноградную кислоту в спирт или молочную кислоту. Организмы, нуждающиеся в кислороде (аэробы), превращают пировиноградную кислоту в воду и углекислоту. Для этого требуется участие ряда растительных кислот в циклических сериях реакций, известных как цикл лимонной кислоты, или цикл Кребса (в честь открывшего его).[ ...]