Снова о трипсах. Когда погодой объясняется 78% изменений численности, это не значит, что процессы, зависящие от плотности, не имеют значения.[ ...]
Следовательно, Андреварта и Берч «отвергали традиционное разделение факторов среды на биотические и абиотические, а также на зависящие и не зависящие от плотности, исходя из того, что такое деление неточно и не дает необходимой базы для обсуждения проблем популяционной экологии» (Andrewartha, Birch, 1960).[ ...]
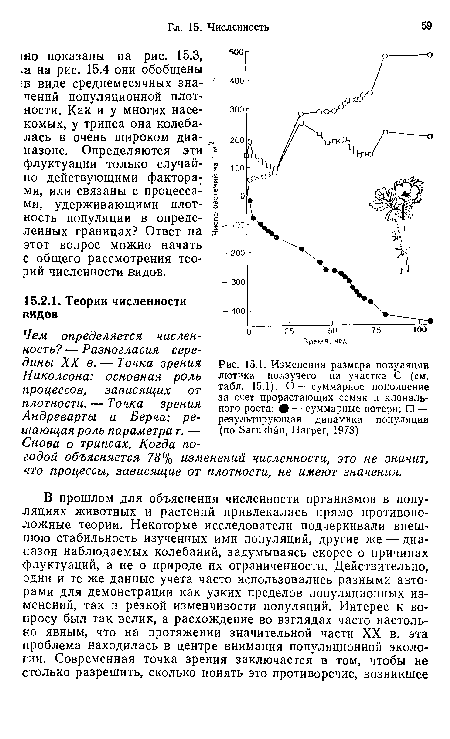
Точку зрения этих авторов можно пояснить, подробнее рассмотрев пример, которому они придавали большое значение (см. рис. 15.3 и 15.4). Уже говорилось, что в ходе учета популяции Thrips imaginis значения численности регистрировались в течение 81 месяца подряд. В течение следующих семи лет оценки проводились лишь весной и в начале лета. Кроме того, на протяжении всего этого периода в данном месте отмечались температура и количество выпадающих осадков. Данные учетов были проанализированы с помощью множественного регрессионного анализа (см., например, Poole, 1978) для установления доли межгодовой популяционной изменчивости, «объясняемой» колебаниями погодных условий. При этом ежегодный максимум обилия популяции рассчитывали как среднее значение логарифма численности за предшествующие максимуму 30 суток. Эти пиковые значения становились в регрессионном анализе зависимой переменной (iogF).[ ...]
Значения Т стали первой независимой переменной (х ) регрессии. Вторая независимая переменная (Х2) представляла собой суммарную величину осадков в сентябре и октябре (весна южного полушария). Независимой переменной хг была суточная «эффективная температура» в сентябре и октябре. Было признано, что размер перезимовавшей популяции мог зависеть от условий прошедшего года. Для учета этого была введена еще одна независимая переменная — значение хх в предшествующий год (л:4).[ ...]
На рис. 15.5 представлены эмпирические и рассчитанные по-уравнению регрессии величины. Важнейшая переменная — хх (Р<0,001), второе место занимает х2 ( <0,01); 78% изменчивости популяционного максимума объяснялось регрессией, «построенной исключительно по метеорологическим данным...[ ...]
Это фактически не оставляет никаких шансов найти какую-либо иную постоянно действующую причину изменчивости, поскольку 22% —достаточно небольшой остаток, который можно связать с ошибками при случайном составлении выборки... Все межгодовые колебания максимума численности, следовательно, вполне объяснимы не зависящими от плотности причинами. Не только не удалось обнаружить «зависящих от плотности факторов», но и показано, что никакой возможности для их существования нет» (Andrewartha, Birch, 1954).[ ...]
Рисунки к данной главе:
| Динамика популяции проломника северного на протяжении восьми лет (по Symonides, 1979а) |
 |