Разорванные ареалы и «нунатаки— Постепенное изменение в распространении древесных пород со времени последнего оледенения. •— Современное межледниковое восстановление — последнее цз многих. — Изменения климата в тропиках.[ ...]
Климат изменялся намного быстрее, чем передвигались массивы суши, и в современной картине географического распространения организмов многое отражает определенные фазы восстановления, происходящего после климатических сдвигов. Для объяснения того, как возникли ареалы тех или иных видов, сведения об исторических изменениях климата необходимы в той же мере, что и представления о приспособленности организмов к современным условиям обитания. В роли исторического фактора в таких случаях выступают, в частности, плейстоценовые оледенения.[ ...]
Классические примеры видовых ареалов, для истолкования которых может понадобиться привлечение исторических обстоятельств,— это состоящие из одного или нескольких небольших обособленных участков ареалы ряда холодоустойчивых, зачастую высокоспециализированных цветковых растений арктоаль-пийских флор Северной Америки и севера Европы. Многие из таких растений встречаются в одном-единственном месте, другие населяют два или несколько причудливо разбросанных обособленных «пятен» (т. е. имеют разорванные ареалы — бицент-рические или полицентрические). Типичным примером может служить бицентрическое распространение одноцветкового колокольчика (Campanula uniflora) в Норвегии (рис. 1.3).[ ...]
Согласно одной точке зрения, отдельные части таких ареалов соответствуют областям, пригодным для произрастания растений данного вида и предоставляющим необходимые для них особые, неповторимые условия. Полагают, что диаспоры таких растений попали в эти области, преодолев промежуточные, непригодные для произрастания, пространства. Согласно этой гипотезе, возможно существование и других областей, где эти растения могли бы прочно обосноваться, но куда они еще не попали. Действительно, растения многих видов с необычными ареалами приурочены либо к островкам известковых почв, либо к определенным температурным условиям (см. рис. 1.3), но так бывает не всегда.[ ...]
Впрочем, спор о нунатаках-—это лишь небольшая часть гораздо более широкой дискуссии между теми, кто считает, что современное распространение организмов отражает их соответствие современным условиям существования, и теми, кто считает, что многое в распространении представляет собой наследие прошлого. С усовершенствованием методов поиска, анализа и датирования биологических остатков (особую роль при этом играет анализ ископаемой пыльцы) размах климатических и биотических сдвигов, происходивших на протяжении последних 2 млн. лет (т. е. в плейстоцене), только начинает приоткрываться. Эти новые методы позволяют все чаще и все с большей уверенностью устанавливать, что именно в современной картине географического распространения организмов связано с узкой специализацией по отношению к нынешним условиям их местообитаний, а что являет собой отпечаток прошедшего.[ ...]
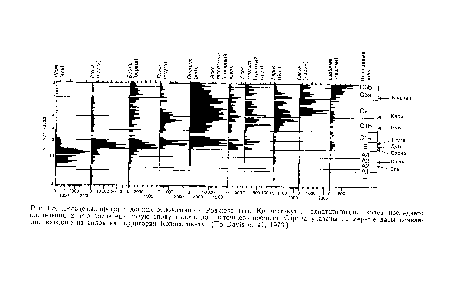
С ростом числа расшифрованных «пыльцевых летописей» стало возможно не только составить представление о динамике растительного покрова в данном месте, но и приступить к картированию перемещений различных видов растений по мере их распространения по материкам. Выясняется, что различные виды продвигались с различными скоростями, и при этом не обязательно в одном и том же направлении. Равновесие растительных сообществ тем временем сдвигалось, состав их изменялся, и изменения эти почти наверняка продолжаются по сей день.[ ...]
Столь же надежными летописями послеледникового расселения животных, связанного с изменениями лесного покрова, мы не располагаем; но по крайней мере не вызывает сомнений то, что многие животные не могли расселяться быстрее тех деревьев, частями которых они питались. Вероятно, некоторые животные все еще пребывают в погоне за отошедшим к северу лесом.[ ...]
Видовой состав лесов умеренных широт Северного полушария непрерывно изменялся в течение прошедших 10 000 лет и продолжает изменяться в настоящее время. Скорость этого изменения, по-видимому, лимитируется скоростями распространения основных лесообразующих пород. Согласно замечанию Девиса (D avis, 1976), то обстоятельство, что «...даже в конце голоценового (т. е. современного) межледниковья заселение освобожденных ледником пространств лесными древовидными растениями все еще происходит, свидетельствует о том, что продолжительность типичной межледниковой эпохи слишком мала для достижения флористического равновесия».[ ...]
По полноте сведений об изменениях климата тропики намного уступают умеренной зоне. По этой причине возникает соблазн предположить, что в те времена, когда умеренные широты находились во власти драматических перемен климата и ледниковых вторжений, тропики .пребывали в том же состоянии, что и ныне. Такое предположение почти наверняка ошибочно. Напротив, складывается картина изменений растительного покрова, параллельных тем, что происходили в умеренном поясе: в более теплые и влажные периоды площадь тропических лесов возрастала, а в периоды более прохладные и засушливые, когда преобладала саванна, — сокращалась. В современном распространении как растений, так и животных содержатся указания на то, какое положение занимали некогда эти «острова тропического леса, окруженные морем саванны» (рис. 1.6).[ ...]
Рисунки к данной главе:
Вернуться к оглавлению