Генеральная закономерность воздействия температуры на живые организмы выражается действием ее на скорость обменных процессов. Согласно общему для всех химических реакций правилу Вант-Гоффа, повышение температуры ведет к пропорциональному возрастанию скорости реакции. Разница заключается в том, что в живом организме химические процессы всегда идут с участием сложных ферментных систем, активность которых в свою очередь зависит от температуры. В результате ферментативного катализа возрастает скорость биохимических реакций и количественно меняется ее зависимость от внешней температуры.[ ...]
Коэффициент температурного ускорения £?ю, для большинства химических реакций абиотического характера равный 2—3, в реакциях живых систем колеблется в довольно широких пределах даже для одних и тех же процессов, протекающих в разных диапазонах температур. Это объясняется тем, что скорость ферментативных реакций нег является линейной функцией температуры. Так, у тропических растений при температуре менее 10°С коэффициент О о приблизительно равен 3, но существенно уменьшается при возрастании температуры выше 25—30°С. У колорадского жука потребление кислорода в диапазоне 10—30°С характеризуется величиной Ою — 2,46, а при температуре 20—30°С Ох о = 1,8. Зависимость метаболизма рыб и многих других водных животных от температуры выражается в изменении величины 010 от 10,9 до 2,2 в диапазоне температур от 0 до 30°С.[ ...]
Тптгцщнм пори тптг Объективная зависимость скорости реакций слг температуры ухе исходно определяет, что жизненные функции могут протекать яинп» в определенном интервале температур. Имеется ряд дополнительных обстоятельств, определяющих температурные порога, выше и ниже которых жизнь невозможна Видовая специфика ферментных систем приводит к тому, что эти пороги неодинаковы для разных видов живых организмов.[ ...]
У более сложно организованных растений и животных тепловая гибель обычно наступает при более низких температурах. Основная причина ее —рассогласование обменных процессов, вызванное разным значением £?ю для разных реакций. У животных большое значение имеют нарушения деятельности нервной системы и ее регуляторных функций. Поэтому у большинства животных тепловая гибель наступает раньше, чем начинают коагулировать белки,—при температуре тела порядка 42—43°С. Растения, обитающие в степях, саваннах, пустынях, выдерживают нагревание до 50—60°С.[ ...]
Нижний температурный порог жизни. Нарушения метаболических и регуляторных процессов наступают и при очень низких температурах. Дисгармония функций в целом организме определяется, как и при гипертермии, разной величиной 0о отдельных реакций. Например, нарушения деятельности сердца при слабом охлаждении проявляются в ритме сокращений и сократимости сердечной мышцы, а при более сильном —в ее проводимости и возбудимости. При одном и том же снижении температуры удлинение периода диастолы выражено сильнее, чем систолы. В почках млекопитающих канальцевая реабсорбция затормаживается уже при температуре тела 23—20°С, а клубочковая фильтрация —только при 19°С, что нарушает выделительную функцию уже при относительно высокой температуре. У насекомых охлаждение подавляет механизмы, обеспечивающие приток кислорода к клеткам, сильнее, чем интенсивность клеточного дыхания. Переваривание пищи в кишечнике пчел тормозится охлаждением в большей степени, чем потребление глюкозы тканями.[ ...]
Нехолодостойкие растения (обитатели влажных тропических лесов, водоросли теплых морей) погибают при температуре несколько выше 0°С из-за инактивации ферментов и нарушения рада метаболических процессов.[ ...]
Важное значение в определении нижнего температурного порога жизни имеют структурные изменения в клетках и тканях, связанные с замерзанием внеклеточной и внутриклеточной жидкостей. При образовании кристаллов льда механически повреждаются ткани, что часто служит непосредственной причиной Холодовой гибели. Кроме того, образование льда нарушает обменные процессы: обезвоживание цитоплазмы влечет за собой повышение концентрации солей, нарушение осмотического равновесия и денатурацию белков. Для многих животных именно нарушения метаболизма вызывают холодовую гибель. Среди растений морозоустойчивые формы выдерживают полное зимнее промерзание, что определяется сезонной перестройкой ультраструктуры клеток, направленной на устойчивость их к обезвоживанию. Сухие . семена выдерживают охлаждение практически до абсолютного нуля.[ ...]
В пределах изменений температуры от верхнего до нижнего порогов жизни реализуется закономерное влияние ее на жизненные процессы, отраженное в правиле Вант-Гоффа.[ ...]
Принципы теплообмена организма. Рассмотренные закономерности отражают зависимость обменных реакций от температуры тела. Последняя же в большинстве случаев не идентична температуре среды; она устанавливается в результате баланса тепла между организмом и внешней средой. Постоянно происходящий обмен тепла (теплообмен) организма со средой зависит от ряда факторов и в принципе складывается из двух противоположных процессов: притока тепла и отдачи его во внешнюю среду. Поступление тепла в организм из внешней среды идет путем теплопроводности и радиации; кроме того, в любом живом организме продуцируется эндогенное тепло как результат всех метаболических реакций. Отдача тепла во внешнюю среду осуществляется также проведением и радиацией; кроме того, значительное количество тепла расходуется организмом в процессе жизнедеятельности путем испарения влаги (скрытая теплота испарения при 22°С составляет 2443,5 Дж/г, или 584 кал/г). Баланс этих двух процессов и определяет собой температуру тела, т. е. тепловую среду биохимических и физиологических реакций, протекающих в организме.[ ...]
Относительная роль перечисленных составляющих теплообмена неодинакова у разных форм живых организмов. По принципиальным особенностям теплообмена различают две крупные экологические группы организмов: пойкилотермные и гомойотермные.[ ...]
Рисунки к данной главе:
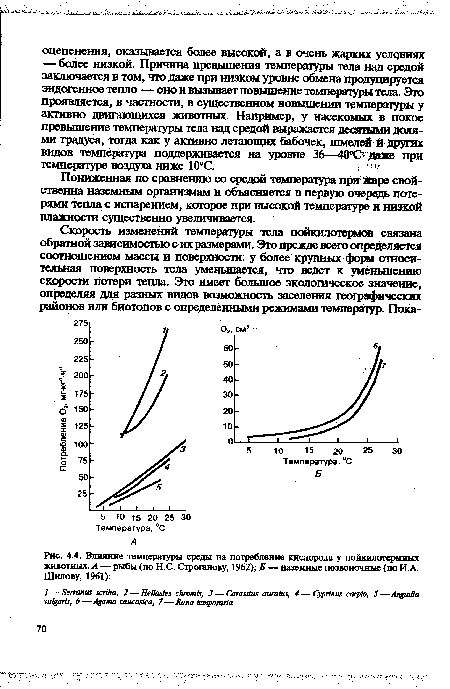
| Влияние температуры среды на потребление кислорода у пойкнлотермиых животных. А — рыбы (по Н.С. Строганову, 1962); Б — наземные позвоночные (по И А. Шилову, 1961) |
 |
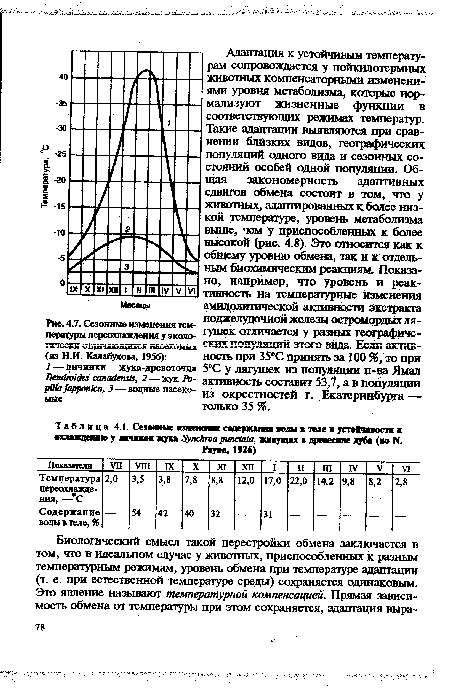
| Сезонные изменения температуры переохлаждения у экологически отличающихся насекомых (из Н.И. Калабухова, 19S6) |
 |
| Пример практически полной и хвостового^ отделов определялась температурной компенсации |
 |
| Потребление кислорода при разной темпе- 11НИ. V г.™ |
 |