Исследование детального пространственного расположения атомов в кристаллической структуре целлюлозы можно разбить на два этапа: первый — определение геометрической конформации изолированной цепи; второй — установление взаимного расположения соседних цепей. Некоторые полиморфные формы целлюлозы могут отличаться друг от друга в обоих отношениях (разд. I, В.2). Поскольку конформация и размеры молекул в растворе (разд. II, Ж) не обязательно такие же, как и в твердом состоянии, изучение кристаллической структуры соответствующих олигосахаридов представляет важный источник для понимания стереохимии целлюлозы. При перенесении закономерностей, полученных для пизкомолеку-лярных соединений, на полимеры следует соблюдать осторожность, однако структурный анализ полимеров, макромолекулы которых построены из остатков p-d-глюкозы, позволит уточнить действительную структуру высокоупорядоченной целлюлозы.[ ...]
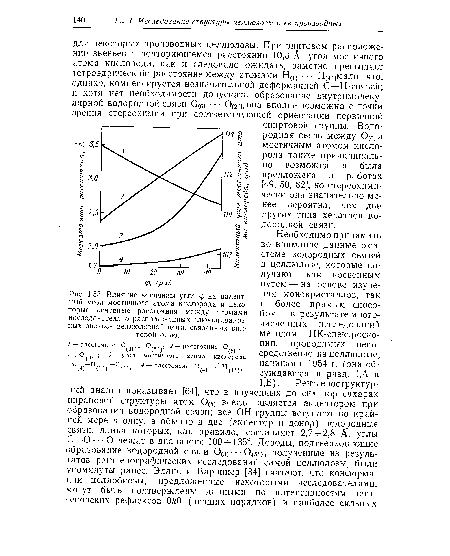
Среди сильных ЛО/-рефлексов гораздо более интенсивными являются те, для которых сумма /г + / — четное число. Это указывает на то, что в пределах ограничений, связанных с неоднозначностью отнесения рефлексов, основные массы рассеивающего вещества разделены расстояниями х = Ч2, 2= /2- Такое разделение на центральные и угловые цепи (центральные, конечно, не в смысле наличия кристаллографического центра симметрии), вытекающее из наличия очень интенсивных 002, 101 и 101 рефлексов, было принято большинством исследователей. Для остальных рентгеновских рефлексов сравнение наблюдаемых и вычисленных интегральных интенсивностей проводилось весьма редко. Наиболее известным исключением является структура Майера и Миша [8], предложенная до того, как был выполнен рентгеноструктурный анализ соответствующих сахаров и проведено исследование целлюлозы методом ИК-спектроскопии. Она оказалась неприемлемой [45] даже при условии, что углы 0 и ф равны 0° (обозначения см. у Джонса [45]). Такой же неприемлемой является предложенная в работе [49] модель целлюлозы I. В этой модели допускаются плоское кольцо, абсолютно невероятные значения углов связей у атома кислорода [77], несовершенная система водородных связей и придается слишком большое значение плоскостям 002 [45]. Уместно напомнить, что интерференции от плоскостей 200, 201, 102, 201 и 102 попадают в область, обычно приписываемую плоскости 002.[ ...]
Фрей-Висслинг [95] пытался преобразовать антипараллельную структуру Майера и Миша в структуру с водородными связями в плоскости 101, а не 002, на том основании, что эти плоскости морфологически более важны, чем плоскость 002. Это предположение, однако, не всегда справедливо [96] и не может служить надежным основанием для предлагаемой структуры. Проверка этой структуры на пространственной модели [97] показала, что ие связанные водородными связями атомы 0(<5) • • • О (з») соседних угловых цепей расположены на меньшем расстоянии, чем связанные водородной связью атом кислорода первичной спиртовой группы (0(6)) угловой цеии и мостичный атом кислорода центральной цеии. Таким образом, для структуры, основанной на маловероятной конформации цеии, иолучают неудовлетворительные данные о длинах и направлениях водородных связей; кроме того, в этой модели атом кислорода 0(2) не участвует в образовании водородной связи. Если принять более вероятную конформацию цеии с хелатной связью О(з) • • • 0(5 ), то следует допустить образование межмолеку-лярной связи между 0(2) и 0(б"), так что наряду со связями, предложенными Фрей-Висслингом, формально можно постулировать систему водородных связей во всех трех плоскостях 002, 101 и 101. При этом, однако, остаются невыясненными некоторые стереохи-мические детали и непонятно, каким образом можно достигнуть совпадения с наблюдаемыми интенсивностями четных ОйО-реф-лексов. Другой вариант [94] структуры Фрей-Висслинга хорошо согласуется с наличием заметного параллельного дихроизма для деформационных колебаний СН2-груип. Однако при построении пространственных моделей с параметрами в области 8 = 25°, ф = = 34° и значением r-сдвига (смещение антипараллельных цепей вдоль осп 6) около ь/з проявился ряд недостатков этой модели [97]: более короткие, чем это следует из частот соответствующих валентных колебаний, растояния 0(2> (угловая цепь) • • ■ О (б") (угловая цепь) и 0(6) (угловая цепь) ■■■0(i) (центральная цепь); заметное отклонение угла 0(й)—Н(6) (угловая цепь) •••0
Имеется и другое расхождение между вычисленными и измеренными интенсивностями [99]: в то время как для регулярной структуры отношение между суммарной интенсивностью экваториальных рефлексов и общей интенсивностью слоевых линий более высокого порядка (исключая меридиональные рефлексы) должно составлять приблизительно 1:1, наблюдаемое отношение равно примерно 5: 1. Таким образом, учитывая результаты спектроскопических и стереохимических исследований, не удается согласовать рентгеноструктурные данные с какой-либо общепринятой кристаллической структурой.[ ...]
Таким образом, различия между разновидностями целлюлоз III и IV согласуются с экспериментально определенными различиями между производными основных модификаций целлюлозы I и II. Эти различия могут быть связаны с конформацией цепей, их полярностью, величиной r-сдвига вдоль оси b или с взаимным, расположением цепей в перпендикулярных направлениях.[ ...]
Рисунки к данной главе:
| Плоская система водородных связей (слой 002), предложенная для целлюлозы I (первый вариант). |
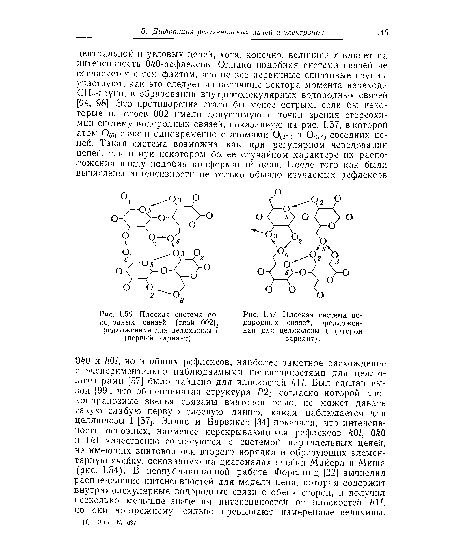 |
| Плоская система водородных связей, предложенная для целлюлозы 1 (второй вариант). |
|
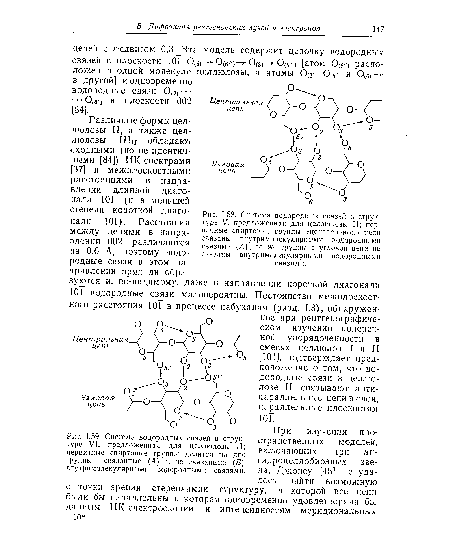
| Система водородных связей в структуре VI, предложенная для целлюлозы II; первичные спиртовые группы делятся на две группы — связанные (А) и не связанные (Б) внутримолекулярными водородными связями. |
 |