Любой фактор — неважно, лимитирующий шГи благоприятный — может быть: 1) не зависящим от плотности (определяющим плотность), если его влияние не зависит от величины популяции, или 2) зависящим от плотности (определяемым плотностью), если его действие на популяцию есть функция ее плотности. Влияние факторов второй группы обычно усиливается по мере приближения к верхнему пределу плотности, но оно может подчиняться также обратной зависимости: с увеличением плотности влияние уменьшается. Факторы, влияние которых находится в прямой зависимости от плотности популяции, действуют подобно регулятору числа оборотов на двигателе; их рассматривают как один из главных механизмов, предотвращающих перенаселение и способствующих установлению устойчивого равновесия. Действие климатических факторов часто (но отнюдь не всегда) не зависит от плотности, тогда как действие биотических факторов (конкуренция, паразиты, патогенные влияния и т. д.) часто (но опять-таки не всегда) зависит от нее.[ ...]
Приступая к интенсивным исследованиям конкретных популяций, мы прежде всего не должны забывать об интеграции на уровне сообщества и экосистемы, поскольку (как подчеркивалось в гл. 2) часть (популяция) никогда нельзя понять без рассмотрения целого (экосистемы) и наоборот. В тех случаях, когда к какому-либо трофическому уровню относится много видов (например, растительноядные насекомые в лесу), популяционная экология любого отдельно взятого вида не будет иметь смысла, если мы не будем знать, что делают в это же самое время его «коллеги».[ ...]
Следовательно, регулирование в популяции в конечном счете есть функция той экосистемы, частью которой эта популяция является. Сделанное нами противопоставление «физически регулируемых» и «биологически регулируемых» экосистем произвольно, но, хотя в данном случае мы пользуемся упрощенной «моделью», такой подход вполне оправдан, особенно в связи с тем, что в результате деятельности человека для биологических систем создаются все более неблагоприятные условия, в которых они не способны ни к самоподдержанию, ни к саморегулированию. Природные экосистемы умеренной зоны, лучше всего изученные с экологической точки зрения, с точки зрения значимости для них физических и биотических регуляторов занимают промежуточное положение. Милн (1957 и 1962) подчеркивает, что механизмы не зависящей и зависящей от плотности регуляции взаимодействуют, и указывает, что последние редко бывают «совершенными» (как при логистическом росте), в частности это обусловлено тем, что лимитирующие влияния при одной плотности более эффективны, чем при другой. В полевых исследованиях отмечалось, что какой-либо регулирующий фактор, например хищник, может быть весьма эффективным при низкой плотности добычи и совсем неэффективным при ее высокой плотности, когда изменение численности хищника наступает лишь с некоторым лаг-периодом после изменения плотности добычи. Холдинг (1961) приходит к выводу, что у насекомых при низкой плотности добычи хищничество с увеличением плотности добычи возрастает, а при высокой плотности уменьшается (т. е. находится в обратно пропорциональной зависимости от плотности).[ ...]
Строго говоря, эффект следует считать не зависящим от плотности только в том случае, если влияние, которому подвергается популяция, постоянно безотносительно к ее размерам. Например, если определенному числу охотников позволяется отстрелять за сезон двух оленей и они охотятся до тех пор, пока не добудут их, то каждый год независимо от общей численности оленей будет изыматься одно и то же их количество. Однако часто говорят, что тот или иной фактор не зависит от плотности, если при изменении плотности популяции его действие остается тем же в процентном выражении (например, если бы каждый год изымалось 10% оленей, в этом случае, разумеется, их общее число при разной плотности было бы разным!).[ ...]
Наиболее существенно в нашей общей теории утверждение, что «популяции имеют выраженную тенденцию эволюционировать таким образом, чтобы достигнуть состояния саморегуляции». В предыдущем разделе мы описали, как физиологические и генетические сдвиги или смена экотипов во времени могут приводить к затуханию осцилляций и ускорять возвращение к более низким уровням плотности. В 60-е годы были получены данные, показывающие, что дело вовсе не в тенденции популяций избегать гибельных отклонений в обе стороны от уровней насыщения; они эволюционируют так, что регуляция их плотности осуществляется на значительно более низкой по сравнению с верхней асимптотой емкости местообитания, достигаемой лишь в том случае, если полностью используются ресурсы энергии и пространства. При этом естественный отбор действует в направлении максимального повышения качества среды обитания особи и уменьшает вероятность гибели популяции. Случай саморегуляции популяции освещен в работах Винн-Эдвард-са (1962 и 1965); регуляция на основе межвидовых взаимодействий, осуществляемых как процесс генётической обратной связи, рассматривается Пиментелом (1961 и 1968). Винн-Эдвардс установил существование двух механизмов стабилизации плотности при ее величинах, лежащих ниже уровня насыщения; 1) территориальное поведение, представляющее собой форму внутривидовой конкуренции, которая, как мы покажем в разд. 15, лимитирует рост популяции посредством регулирования «землепользования»; 2) групповое поведение, выражающееся, например, в «порядке клевания», «половом доминировании» и других поведенческих особенностях, которые нам предстоит обсудить в гл. 8. Что касается генетической обратной связи, то она служит тем механизмом, посредством которого тесно связанные популяции «навязывают» свои требования друг другу. Говоря словами Пиментела (1961), в системе растительноядное животное — растение плотность животных играет для растения роль фактора отбора, этот отбор влияет на генетическую конституцию растений, а генетическая конституция растений в свою очередь влияет на плотность животных. Взаимное влияние друг на друга организмов, входящих в пищейую цепь, совместно с генетическими механизмами обратной связи служит основой эволюции и регуляции популяций. Такого рода «сопряженная эволюция» экологически взаимосвязанных популяций будет снова, но более подробно рассматриваться в гл. 9, разд. 5.[ ...]
Наиболее характерные примеры взаимодействий, не зависящих от плотности, — сильные бури, внезапные падения температуры и другие резкие изменения физических условий. Исследуя в течение трех лет улитку Астаеа, живущую на скалах в приливно-отливной зоне, Фрэнк (1965) обнаружил, что большая часть изменений величины популяции зависит от плотности; исключение составляют только случаи резкого увеличения смертности в годы с суровой зимой, когда внезапно откалываются целые куски скал, так что из популяции изымаются все имеющиеся на них улитки независимо от их числа. Нами уже был рассмотрен хороший случай «совершенной» зависимости от плотности у дрожжей (фиг. 83). Вероятность выявления линейной зависимости от плотности при межвидовых взаимодействиях меньше. Проведя тщательное исследование орехотворки, Варли (1947) показал, что влияние ее главного паразита, насекомого Eurytoma curta, может зависеть от плотности, поскольку в условиях, когда популяция хозяина велика, от этого паразита погибает большее (как в процентном, так и в абсолютном выражении) число особей хозяина (табл. 27).[ ...]
Однако в другом исследовании Варли и Эдвардс (1957) сообщили, что, когда «радиус действия» мал, как в случае паразитической осы МогтотеИа, действие паразита на двукрылых насекомых-хозяев не обязательно зависит от плотности. Таким образом, важную роль могут играть различия в поведении. Холлинг (1965, 1966) предложил серию изящных математических моделей, в которые введены поведенческие характеристики; эти модели позволяют прогнозировать эффективность регулирующего влияния данного насекомого-паразита на насекомое-хозяина при различных уровнях плотности. Подробнее об этом см. в гл. 10.[ ...]
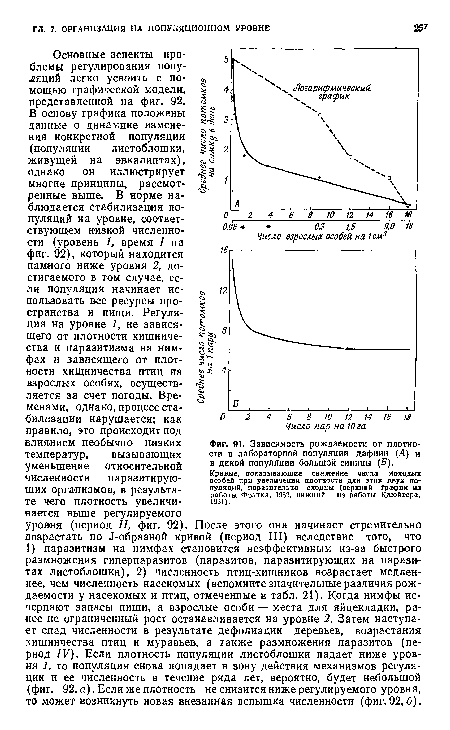
То, что рождаемость и смертность могут изменяться в зависимости от плотности, показывают два примера, представленные на фиг. 91, один из них относится к лабораторной, а другой — к природной популяции. Видно, что с увеличением плотности в лабораторных культурах дафнии и в диких популяциях большой синицы продукция яиц и молодых особей на самку уменьшается. В обоих случаях связь по существу носит экспоненциальный характер (и может быть выражена тем же основным уравнением, которое используется для роста популяций).[ ...]
Пример генетической обратной связи для случая регулирования популяций за счет межвидовых взаимосвязей приведен на фиг. 105.[ ...]
Основные аспекты проблемы регулирования популяций легко усвоить с помощью графической модели, представленной на фиг. 92.[ ...]
Рисунки к данной главе:
Вернуться к оглавлению