| Молекулярные диаграммы НАДН и НАД [12]. |
![Молекулярные диаграммы НАДН и НАД [12].](/static/pngsmall/827884936.png) |
Далее
| Схематическое изображение состояния равновесия между Р-й М-спиральными формами НАДН в водном растворе [47]. |
![Схематическое изображение состояния равновесия между Р-й М-спиральными формами НАДН в водном растворе [47].](/static/pngsmall/827884954.png) |
Далее
| Функционирование имидазольного кольца гистидина в активном центре лактатдегидрогеназы [53]. |
![Функционирование имидазольного кольца гистидина в активном центре лактатдегидрогеназы [53].](/static/pngsmall/827884978.png) |
Далее
| Функционирование е-аминогруппы лизина в активном центре алкогольдегидрогена-зы [103]. |
![Функционирование е-аминогруппы лизина в активном центре алкогольдегидрогена-зы [103].](/static/pngsmall/827884984.png) |
Далее
| Механизм превращения субстрата и кофер-мента в активном центре алкогольдегидрогеназы [88]. |
![Механизм превращения субстрата и кофер-мента в активном центре алкогольдегидрогеназы [88].](/static/pngsmall/827885028.png) |
Далее
| Расположение остатков цистеина в полипептидной цепи ГАФД различного происхождения [3, 4]. |
![Расположение остатков цистеина в полипептидной цепи ГАФД различного происхождения [3, 4].](/static/pngsmall/827885044.png) |
Далее
| Схематическое изображение распределения ароматических аминокислот в ГАФД свиньи и рака согласно данным Harris с соавторами [3, 4]. |
![Схематическое изображение распределения ароматических аминокислот в ГАФД свиньи и рака согласно данным Harris с соавторами [3, 4].](/static/pngsmall/827885048.png) |
Далее
| Зависимость скорости реакции, катализируемой малатде-гидрогеназой, от концентрации субстрата — малата при различных концентрациях аллостерического ингибитора — НАДН [32]. |
![Зависимость скорости реакции, катализируемой малатде-гидрогеназой, от концентрации субстрата — малата при различных концентрациях аллостерического ингибитора — НАДН [32].](/static/pngsmall/827885092.png) |
Далее
| Зависимость скорости ферментативной реакции (V), катализируемой НАД-зависимой изоцитратдегидрогеназой из брюквы, от концентрации НАД в координатах и-^[НАД] (а) и 1/у-МДНАД] (б) [42]. |
![Зависимость скорости ферментативной реакции (V), катализируемой НАД-зависимой изоцитратдегидрогеназой из брюквы, от концентрации НАД в координатах и-^[НАД] (а) и 1/у-МДНАД] (б) [42].](/static/pngsmall/827885096.png) |
Далее
| Десенсибилизация глутаматдегидрогеназы из печени быка при обработке фтордииитро-бензолом [72]. |
![Десенсибилизация глутаматдегидрогеназы из печени быка при обработке фтордииитро-бензолом [72].](/static/pngsmall/827885114.png) |
Далее
| Связывание НАД дрожжевой Д-глицер альдегид-3-фосфат-дегидрогеназой, прослеженное спектрофотометрическим методом [pH 8,5; 40°) [85]; У —степень насыщения. |
![Связывание НАД дрожжевой Д-глицер альдегид-3-фосфат-дегидрогеназой, прослеженное спектрофотометрическим методом [pH 8,5; 40°) [85]; У —степень насыщения.](/static/pngsmall/827885118.png) |
Далее
| Функции насыщения глутаматдегидрогеназы из печени быка аллостерическим ингибитором — ГТФ при различных концентрациях фермента. |
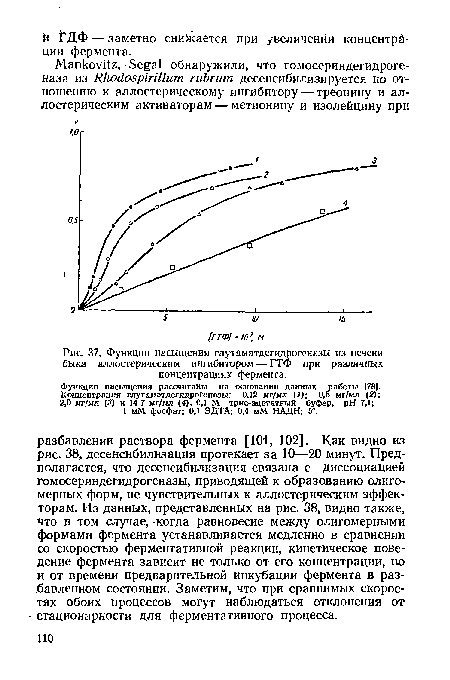 |
Далее
| Зависимость времени удерживания (^тк) от числа изопреноидных звеньев (п). |
 |
Далее
| Сигналы ЭПР митохондрий. |
 |
Далее
| Влияние ингибиторов иа величину индуцированного сукци-натом сигнала, уровень восстановленного (3 и интенсивности дыхания интактных митохондрий (а) и экстрагированных митохондрий с включением 0 (б). |
 |
Далее
| Схема предполагаемого участия семихинонных радикалов в переносе электронов по дыхательной цепи и фосфорилировании. |
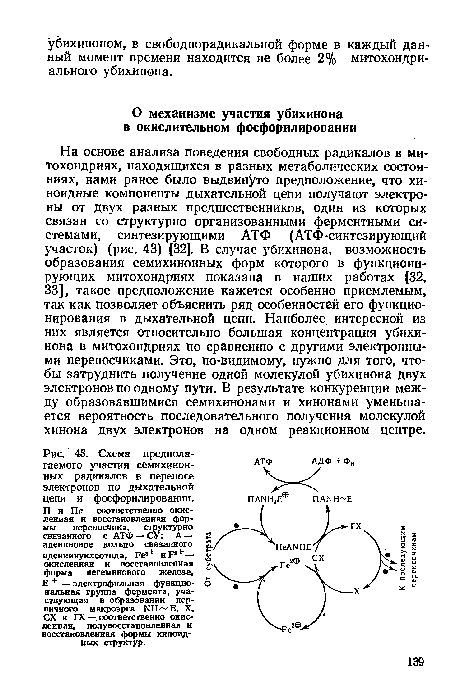 |
Далее
| Спектры поглощен! |
 |
Далее
| Кривые дисперсии оптического вращения ФАД (1); ФМН (2) и АМФ (3). |
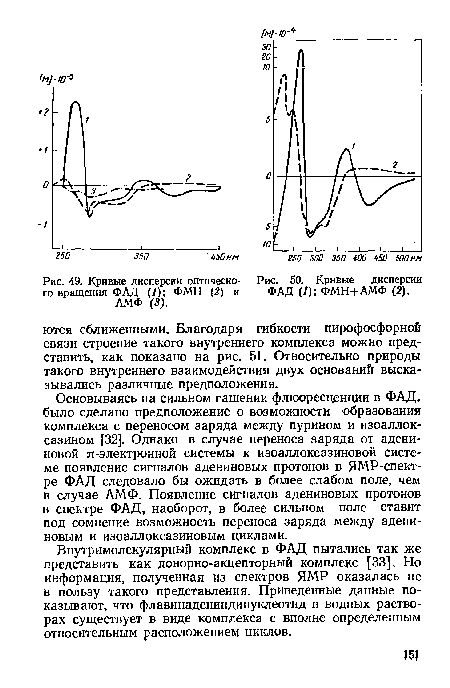 |
Далее
| Кривые дисперсии ФАД (/); ФМН+АМФ (2). |
|
Далее
| Оксидаза ¿-аминокислот. Зависимость скорости реакции от концентрации ¿-лейцина (8) при различных концентрациях кислорода. |
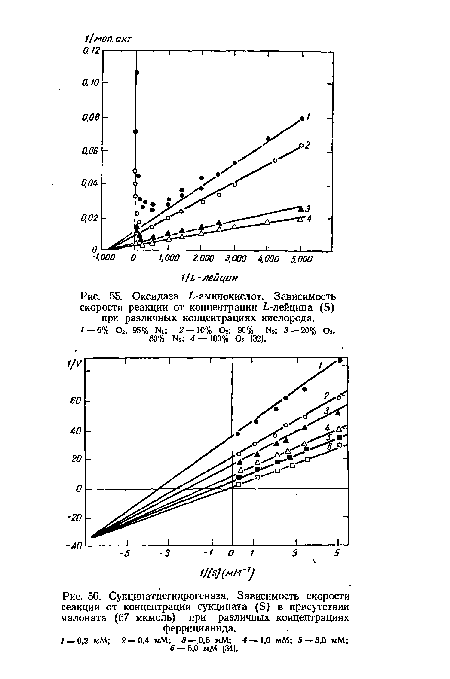 |
Далее
| Спектр сукцинатдегидрогеназы, преинкубированной с сукцинатом [34]. |
![Спектр сукцинатдегидрогеназы, преинкубированной с сукцинатом [34].](/static/pngsmall/827885246.png) |
Далее
| Зависимость скорости реакции от концентрации субстрата при взаимодействии нуклеозиддифосфатсахаров с УДФГ. |
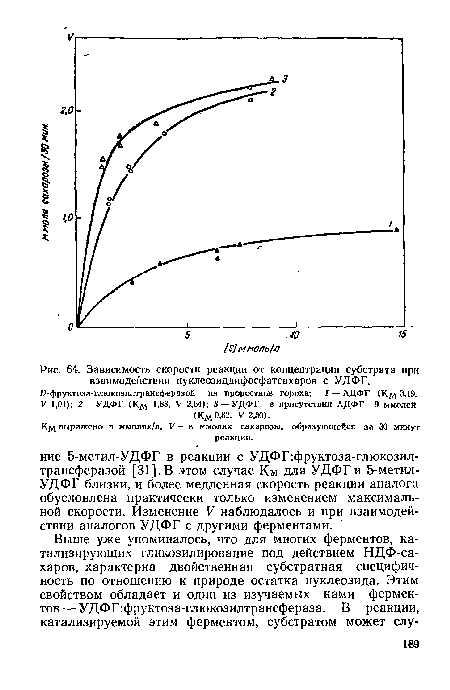 |
Далее
| Связи кофермента с белком в отсутствие субстратов в активной форме фермента, pH 8,0 [см. схему 1 (1)]. |
![Связи кофермента с белком в отсутствие субстратов в активной форме фермента, pH 8,0 [см. схему 1 (1)].](/static/pngsmall/827885330.png) |
Далее
| Фермент-субстратный комплекс 3 [см, схему 1 (3)]. |
![Фермент-субстратный комплекс 3 [см, схему 1 (3)].](/static/pngsmall/827885336.png) |
Далее
| Механизм образования холофермента. |
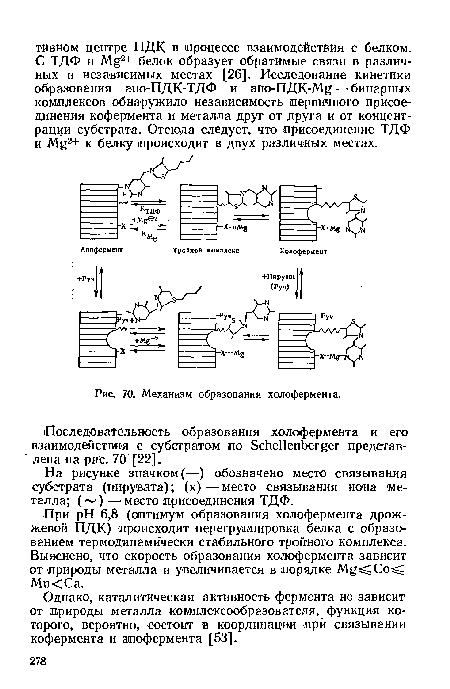 |
Далее
| Взаимное расположение тиамина и 3-иидолилацета-та в молекулярном комплексе. |
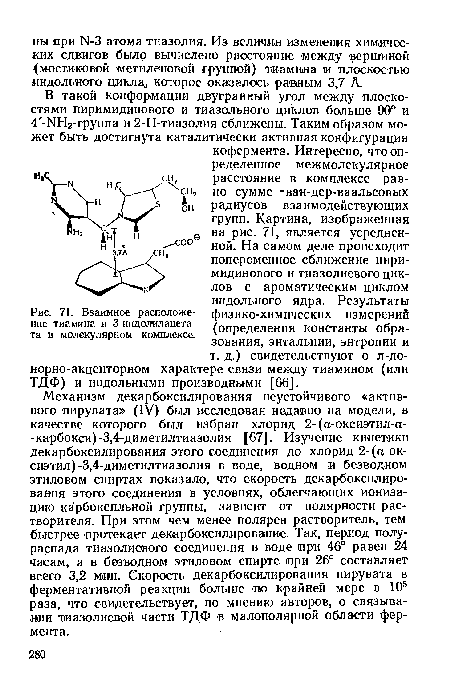 |
Далее
| Структура ДБК-кофермента, полученная на основании данных рентгеноструктурного анализа. |
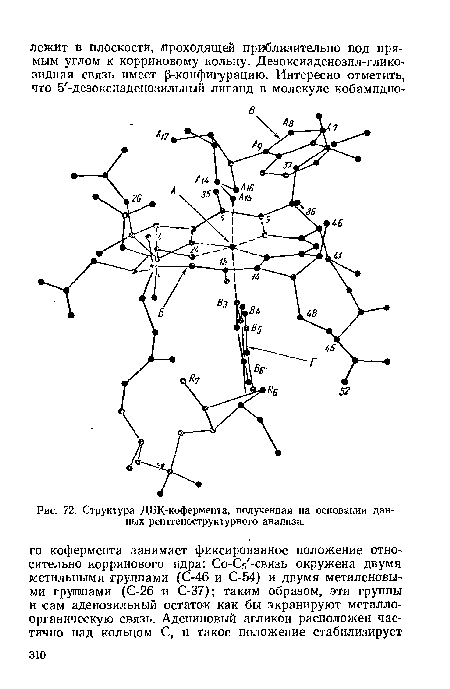 |
Далее
| Спектры поглощения различных корриновых соединений. |
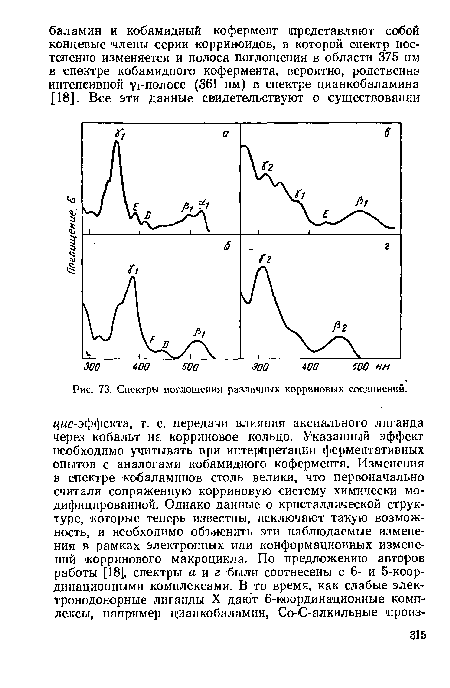 |
Далее
| Два центра связывания кофермента с апоферментом. |
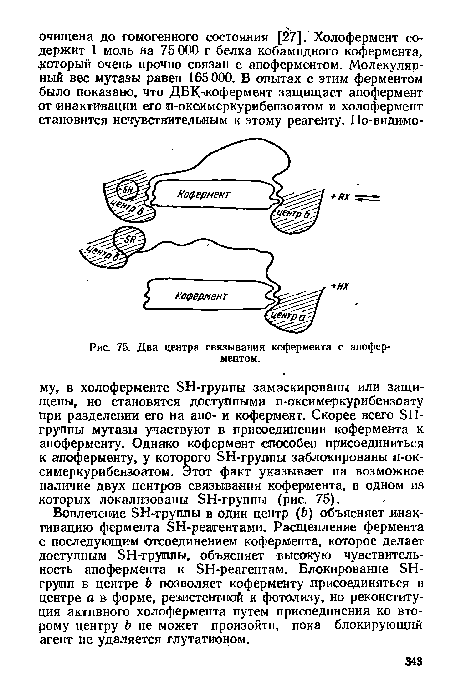 |
Далее
| Постулированный механизм переноса гидридного иона (1> |
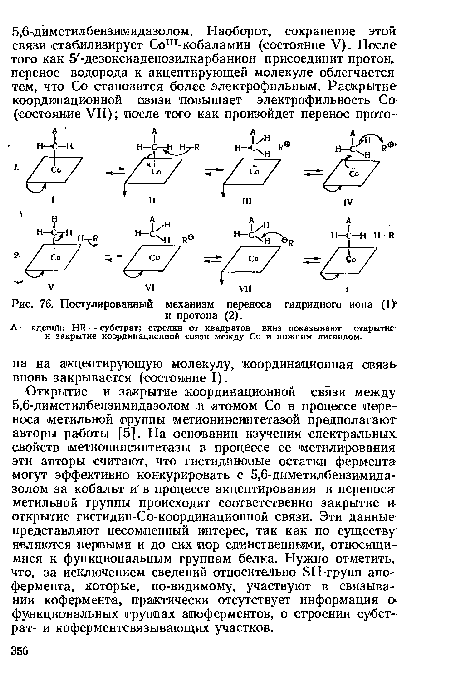 |
Далее
| Зависимость активности альдолазы и ее способности подвергаться триптическому гидролизу от числа сульфгидрильных групп, блокированных п-ХМБ. |
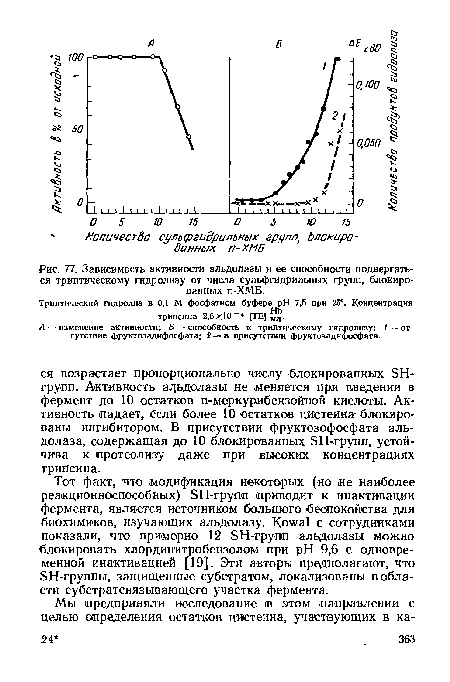 |
Далее
| Образование карбоксиметильных производных альдолазы в ходе реакции алкилирования. Концентрация бромацетата |
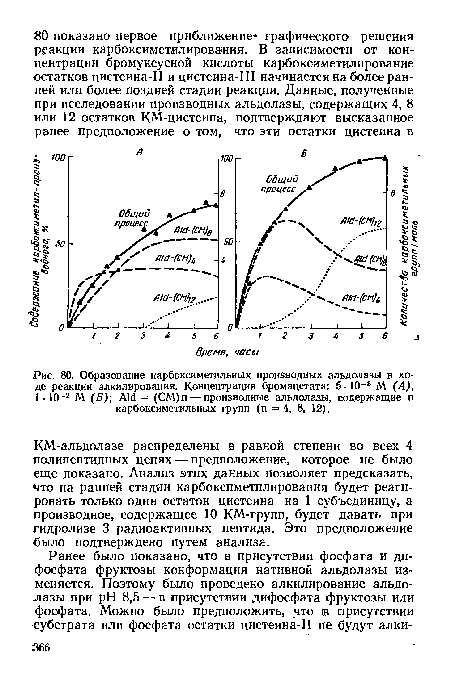 |
Далее
| М (Б) Aid = (СМ)п — производные альдолазы, содержащие п карбоксиметильных групп (п = 4, 8, 12). |
|
Далее
| Реакция алкилирования альдолазы бромацетатом [С14] в присутствии фосфата. Концентрация |
![Реакция алкилирования альдолазы бромацетатом [С14] в присутствии фосфата. Концентрация](/static/pngsmall/827885638.png) |
Далее
| Распределение остатков цистеина в полипептидной цепи альдолазы, полученное на основании анализа бромцианово-го гидролизата. |
 |
Далее