| А. Географическое распространение и степень родства крупных нелетающих птиц можно хотя бы отчасти объяснить дрейфом материков (см. рис. 1.1). Б. Степень родства определялась методом гибридизации ДНК. Ме- тод состоит в следующем. При нагревании двухцепочечная спираль молекулы ■ДНК. распадается на две отдельные цепи. Цепи ДНК различных видов можно затем соединить и вновь разделить с помощью нагревания. Чем выше сходство между цепями, тем более высокая температура (АТ$0Н) необходима для их разделения. Таким образом, эта температура является мерой степени родства двух видов и давности дивергенции. Прежде всего произошла дивергенция линий тинаму и бескилевых птиц. Очередность более поздней дивергенции хорошо согласуется с хронологией раскола Гондваны и последовавшего за ним дрейфа материков (рис. 1.1). Последовательность основных событий такова |  |
А. Географическое распространение и степень родства крупных нелетающих птиц можно хотя бы отчасти объяснить дрейфом материков (см. рис. 1.1). Б. Степень родства определялась методом гибридизации ДНК. Ме- тод состоит в следующем. При нагревании двухцепочечная спираль молекулы ■ДНК. распадается на две отдельные цепи. Цепи ДНК различных видов можно затем соединить и вновь разделить с помощью нагревания. Чем выше сходство между цепями, тем более высокая температура (АТ$0Н) необходима для их разделения. Таким образом, эта температура является мерой степени родства двух видов и давности дивергенции. Прежде всего произошла дивергенция линий тинаму и бескилевых птиц. Очередность более поздней дивергенции хорошо согласуется с хронологией раскола Гондваны и последовавшего за ним дрейфа материков (рис. 1.1). Последовательность основных событий такова
Примерная динамика температуры в течение последовательных циклов оледенения, пришедшихся на последние 400 тыс. лет. Оценки температур получены путем сравнения соотношений изотопов кислорода в ископаемых остатках организмов, извлеченных из кернов донных отложении Карибского моря. Штриховая линия соответствует температуре, имевшей место 10 тыс. лет назад—в начале современного потепления. Кривая показывает, что теплые мажледниковья, подобные современному, по-видимому, случались редко и что в течение большой части прошедших 400 тыс. лет господствовал ледниковый климат. (По Emiliani, 1966; Davis, 1976.)
| Примерная динамика температуры в течение последовательных циклов оледенения, пришедшихся на последние 400 тыс. лет. Оценки температур получены путем сравнения соотношений изотопов кислорода в ископаемых остатках организмов, извлеченных из кернов донных отложении Карибского моря. Штриховая линия соответствует температуре, имевшей место 10 тыс. лет назад—в начале современного потепления. Кривая показывает, что теплые мажледниковья, подобные современному, по-видимому, случались редко и что в течение большой части прошедших 400 тыс. лет господствовал ледниковый климат. (По Emiliani, 1966; Davis, 1976.) |  |
Пыльцевые профили донных отложений оз. Роджерс (шт. Коннектикут), охватывающие конец последнего оледенения и всю послеледниковую эпоху вплоть до настоящего времени. Справа указаны примерные даты появления каждого из видов на территории Коннектикута. (По Davis et al., 1973.)
| Пыльцевые профили донных отложений оз. Роджерс (шт. Коннектикут), охватывающие конец последнего оледенения и всю послеледниковую эпоху вплоть до настоящего времени. Справа указаны примерные даты появления каждого из видов на территории Коннектикута. (По Davis et al., 1973.) | 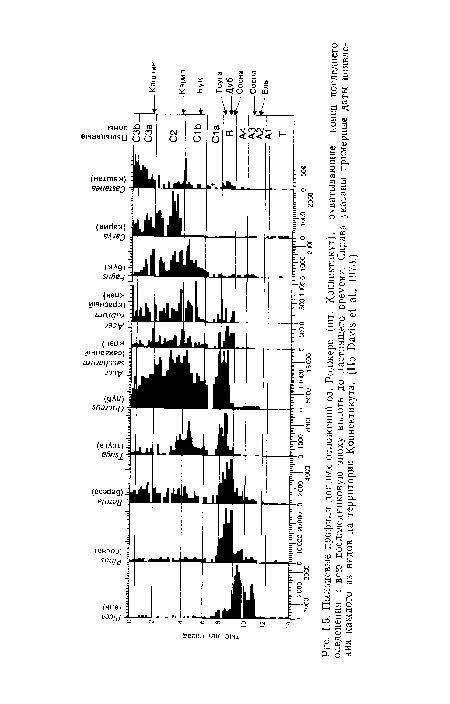 |
А. Примерное распространение тропических лесов в Южной Америке на пике последнего оледенения. Б. Их современное распространение. (По рисункам ЭтЬеНо!^ 1983; Р1е1ои, 1979.)
| А. Примерное распространение тропических лесов в Южной Америке на пике последнего оледенения. Б. Их современное распространение. (По рисункам ЭтЬеНо!^ 1983; Р1е1ои, 1979.) | 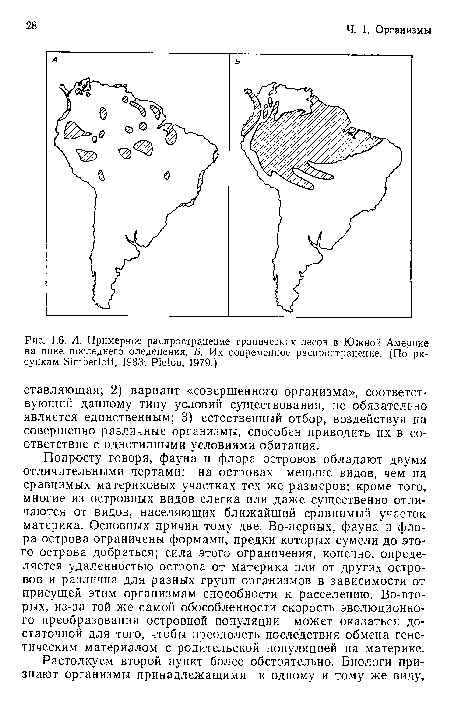 |
А. Дарвиновы вьюрки поедают самую разнообразную пищу и населяют самые разнообразные местообитания; столь же разнообразны по своей форме, несмотря на близкое родство их обладателей, и клювы вьюрков. Б. Распространение различных видов вьюрков на Галапагосских о-вах (включая о. Кокос). На карте указано число видов, встречающихся на каждом из островов. Таблица содержит сведения о расселении каждого из видов по островам; подвиды, входящие в состав каждого данного вида, обозначены различными буквами. (По Lack, 1969а. Названия островов — по Harris, 1973а.) (В головке таблицы (3-я колонка) следует читать
| А. Дарвиновы вьюрки поедают самую разнообразную пищу и населяют самые разнообразные местообитания; столь же разнообразны по своей форме, несмотря на близкое родство их обладателей, и клювы вьюрков. Б. Распространение различных видов вьюрков на Галапагосских о-вах (включая о. Кокос). На карте указано число видов, встречающихся на каждом из островов. Таблица содержит сведения о расселении каждого из видов по островам; подвиды, входящие в состав каждого данного вида, обозначены различными буквами. (По Lack, 1969а. Названия островов — по Harris, 1973а.) (В головке таблицы (3-я колонка) следует читать | 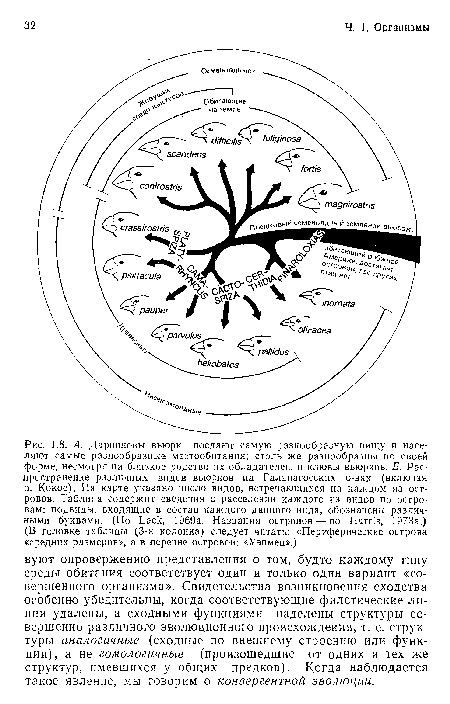 |
Пример конвергентной эволюции
| Пример конвергентной эволюции | 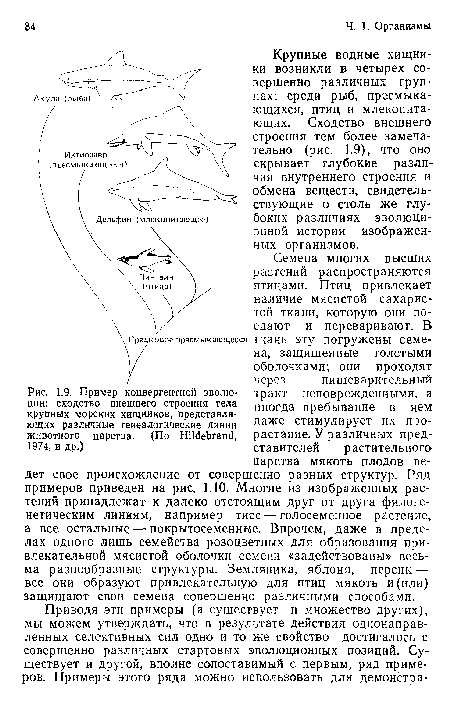 |
Наблюдаемая в различных группах высших растений конвергентная эволюция сочных плодов на основе различных морфологических образований. Гомологичные ткани заштрихованы одинаково.
| Наблюдаемая в различных группах высших растений конвергентная эволюция сочных плодов на основе различных морфологических образований. Гомологичные ткани заштрихованы одинаково. | 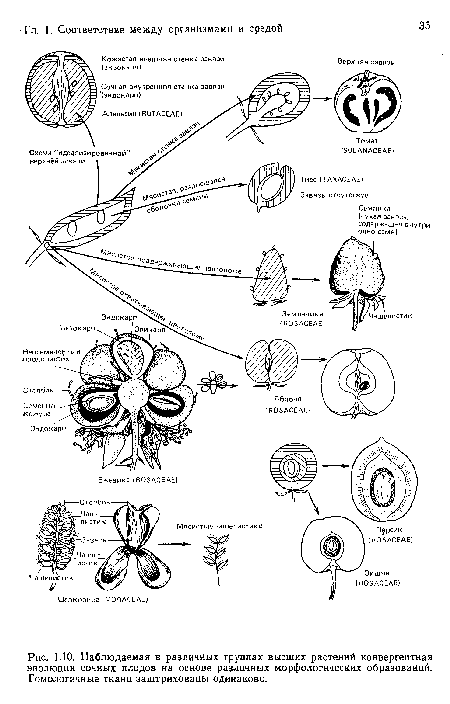 |
Параллельная эволюция сумчатых и плацентарных млекопитающих. Животные сопоставляемых видов напоминают друг друга как по внешнему виду, так и по особенностям образа жизни.
| Параллельная эволюция сумчатых и плацентарных млекопитающих. Животные сопоставляемых видов напоминают друг друга как по внешнему виду, так и по особенностям образа жизни. | 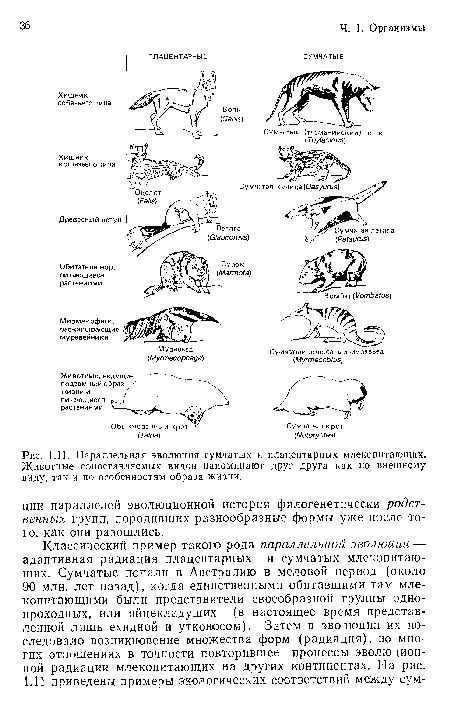 |
Методом Раункиера можно сравнивать целые растительные сообщества. А. Относительное число видов, принадлежащих к той или иной жизненной форме, в случайной выборке из мирового флористического списка (Index Kewensis). Б. Спектр жизненных форм растительности двух тропических островных групп. В. Спектры жизненных форм растительности двух областей умеренного пояса. Г. Спектры жизненных форм растительности двух засушливых местностей. Д. Спектры жизненных форм растительности двух арктических областей.
| Методом Раункиера можно сравнивать целые растительные сообщества. А. Относительное число видов, принадлежащих к той или иной жизненной форме, в случайной выборке из мирового флористического списка (Index Kewensis). Б. Спектр жизненных форм растительности двух тропических островных групп. В. Спектры жизненных форм растительности двух областей умеренного пояса. Г. Спектры жизненных форм растительности двух засушливых местностей. Д. Спектры жизненных форм растительности двух арктических областей. | 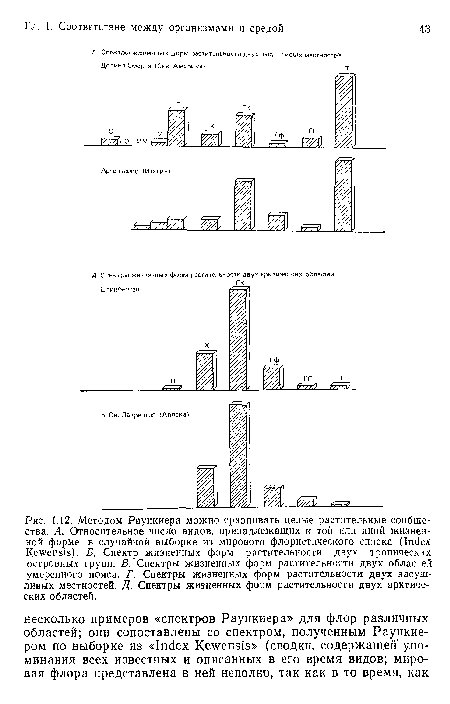 |
Состав пищи некоторых представителен группы антарктических ластоногих, глубина, на которой они добывают пищу, и строение их зубов. (По Ьа У13, 1984.)
| Состав пищи некоторых представителен группы антарктических ластоногих, глубина, на которой они добывают пищу, и строение их зубов. (По Ьа У13, 1984.) | 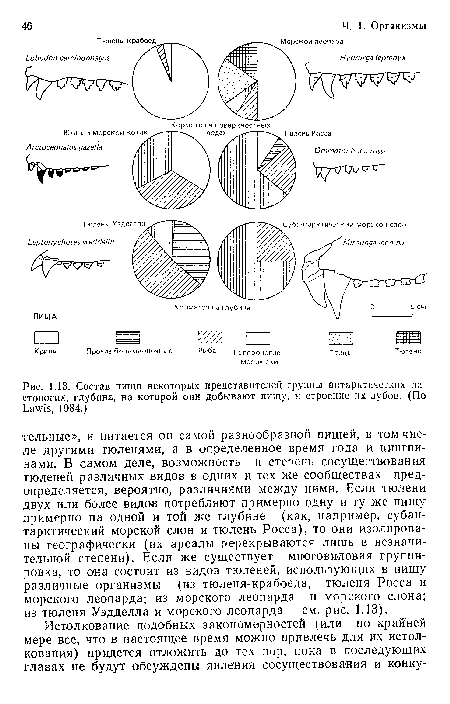 |
Схема расселения слизней Dendroceras по небольшому участку многолетнего пастбища. Контуры дают представление о плотности популяции слизней и разграничивают зоны очень высокой (ОВ), высокой (В), низкой (Н) и очень низкой (ОН) плотности. Указаны также генотипы особей клевера, оказавшихся в узлах накрывающей участок «решетки» проб. Выделены следующие генотипы
| Схема расселения слизней Dendroceras по небольшому участку многолетнего пастбища. Контуры дают представление о плотности популяции слизней и разграничивают зоны очень высокой (ОВ), высокой (В), низкой (Н) и очень низкой (ОН) плотности. Указаны также генотипы особей клевера, оказавшихся в узлах накрывающей участок «решетки» проб. Выделены следующие генотипы | 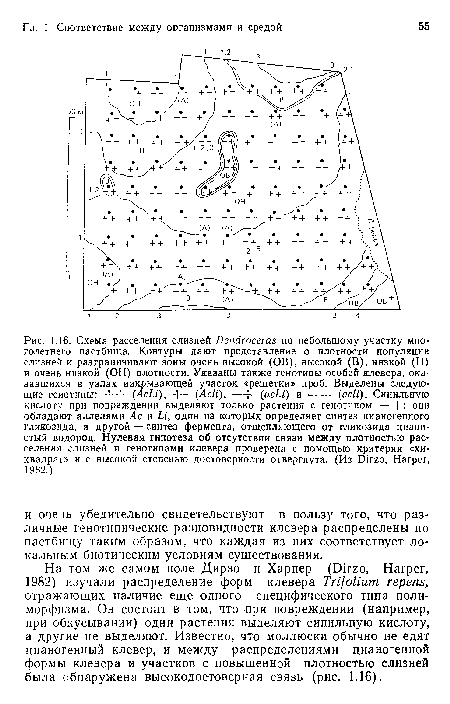 |
Изменчивость количества осадков и непредсказуемость их выпадения. Диаграммы соответствуют трем различным местностям. Каждая из диаграмм отражает многолетнюю динамику количества осадков, выпадающих в течение того месяца, который в соответствующей местности является самым дождливым (в среднем). (Из Stahler, 1960. Данные получены от H. Н. Clayton из Смитсоновского института.)
| Изменчивость количества осадков и непредсказуемость их выпадения. Диаграммы соответствуют трем различным местностям. Каждая из диаграмм отражает многолетнюю динамику количества осадков, выпадающих в течение того месяца, который в соответствующей местности является самым дождливым (в среднем). (Из Stahler, 1960. Данные получены от H. Н. Clayton из Смитсоновского института.) | 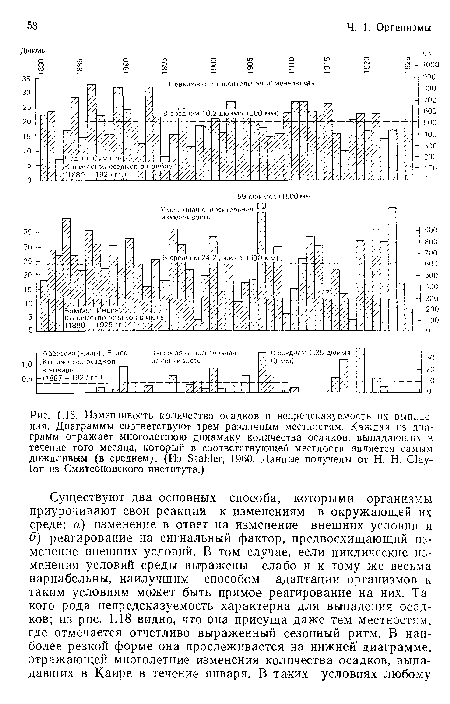 |
Обобщенное графическое изображение зависимости проявлений жизнедеятельности организмов данного вида от степени выраженности внешнего условия. Узкий интервал, в пределах которого особи могут размножаться (И—К), обычно предопределяет те условия, в которых возможно длительное существование вида (впрочем, сколь угодно длительное существование некоторых растений может, по-видимому, поддерживаться исключительно за счет вегетативного роста).
| Обобщенное графическое изображение зависимости проявлений жизнедеятельности организмов данного вида от степени выраженности внешнего условия. Узкий интервал, в пределах которого особи могут размножаться (И—К), обычно предопределяет те условия, в которых возможно длительное существование вида (впрочем, сколь угодно длительное существование некоторых растений может, по-видимому, поддерживаться исключительно за счет вегетативного роста). |  |
Характерные температуры, предпочитаемые рыбами четырех видов. В условиях созданного в лаборатории температурного градиента рыбы собирались в тех местах, где поддерживались эти температуры. Перед началом опыта все рыбы выдерживались при температуре 15°С. (Из Ferguson, 1958.)
| Характерные температуры, предпочитаемые рыбами четырех видов. В условиях созданного в лаборатории температурного градиента рыбы собирались в тех местах, где поддерживались эти температуры. Перед началом опыта все рыбы выдерживались при температуре 15°С. (Из Ferguson, 1958.) |  |
Схематическое изображение путей теплообмена между эктотермиым организмом и разнообразными окружающими его физическими телами. (Из Hainsworth, 1981, по Gates, Porter, 1970.)
| Схематическое изображение путей теплообмена между эктотермиым организмом и разнообразными окружающими его физическими телами. (Из Hainsworth, 1981, по Gates, Porter, 1970.) | 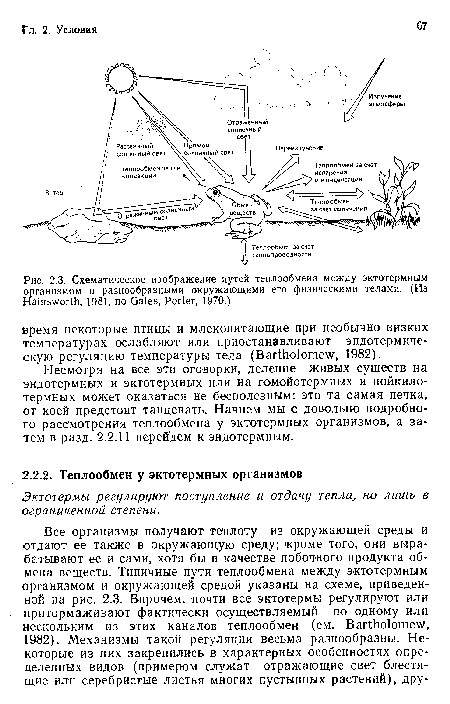 |
Реакция гороха (Pisurn sativum) на температуру зависит от стадии его развития. (По Wang, 1960.)
| Реакция гороха (Pisurn sativum) на температуру зависит от стадии его развития. (По Wang, 1960.) | 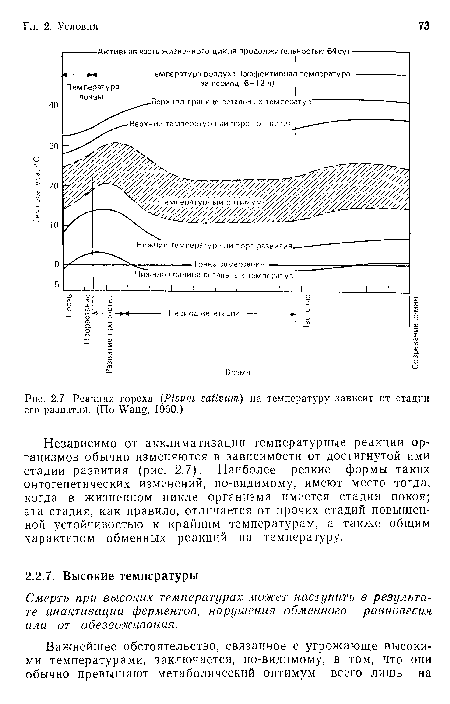 |
Закаливание у растений
| Закаливание у растений |  |
А. Положение Земли на 22 июня
| А. Положение Земли на 22 июня | 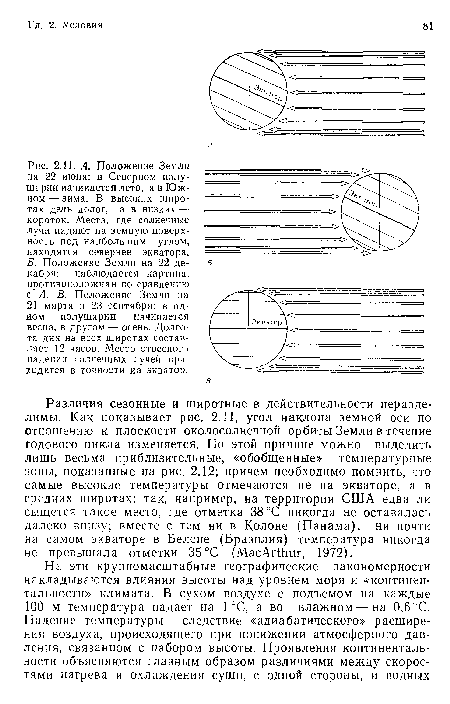 |
Упрощенная схема подразделения поверхности Земли на пять основных климатических зон. (Жирным крестом отмечены Канарские о-ва; см. тскст, с. 85.)
| Упрощенная схема подразделения поверхности Земли на пять основных климатических зон. (Жирным крестом отмечены Канарские о-ва; см. тскст, с. 85.) |  |
Высотная зональность растительности и животного мира горы Кения. (По Сох е1 а1., 1976.)
| Высотная зональность растительности и животного мира горы Кения. (По Сох е1 а1., 1976.) |  |
Экологические ниши. А. Одномерная проекция (на температурную ось). Б. Двумерная проекция (на плоскость «температура—влажность»), В. Трехмерная проекция (например, на подпространство «температура—влажность—скорость течения»).
| Экологические ниши. А. Одномерная проекция (на температурную ось). Б. Двумерная проекция (на плоскость «температура—влажность»), В. Трехмерная проекция (например, на подпространство «температура—влажность—скорость течения»). |  |
А. Одномерные проекции экологических ниш
| А. Одномерные проекции экологических ниш |  |
Смешанный травостой из злаков и ползучего клевера (Trifolium repens)
| Смешанный травостой из злаков и ползучего клевера (Trifolium repens) |  |
Вода в почве. Показано соотношение между тремя характеристиками статуса почвенной влаги
| Вода в почве. Показано соотношение между тремя характеристиками статуса почвенной влаги | 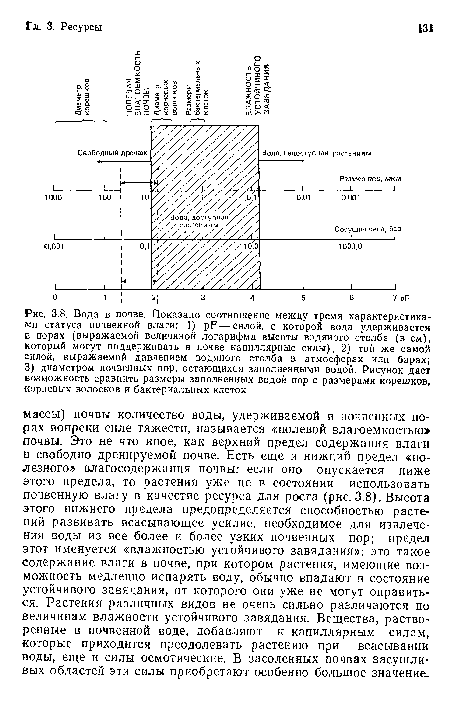 |
Скорость проникновения ограниченного объема воды в толщу сухой суглинистой почвы. (По Russell, 1973.)
| Скорость проникновения ограниченного объема воды в толщу сухой суглинистой почвы. (По Russell, 1973.) | 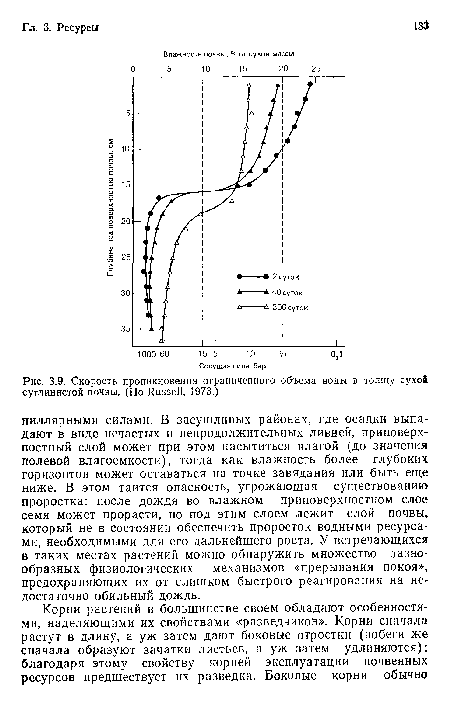 |
Корневая система, образованная растением пшеницы по мере прорастания корней сквозь толщу песчаной почвы, содержавшей прослойку глины. Обратите внимание, как чутко отзывается развитие корня на те сугубо местные условия, в которые он попадает. (Рисунок предоставлен J. V. Lake.)
| Корневая система, образованная растением пшеницы по мере прорастания корней сквозь толщу песчаной почвы, содержавшей прослойку глины. Обратите внимание, как чутко отзывается развитие корня на те сугубо местные условия, в которые он попадает. (Рисунок предоставлен J. V. Lake.) | 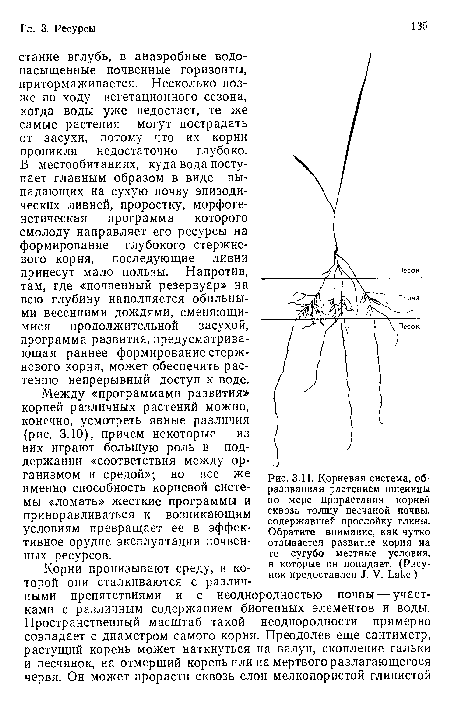 |
Жизненные циклы и сроки протекания последовательных фаз развития растения обыкновенной малины (КиЬиБ Iйаеиз) и некоторых из животных, использующих его в качестве пищевого ресурса (по данным из целого ряда источников)
| Жизненные циклы и сроки протекания последовательных фаз развития растения обыкновенной малины (КиЬиБ Iйаеиз) и некоторых из животных, использующих его в качестве пищевого ресурса (по данным из целого ряда источников) | 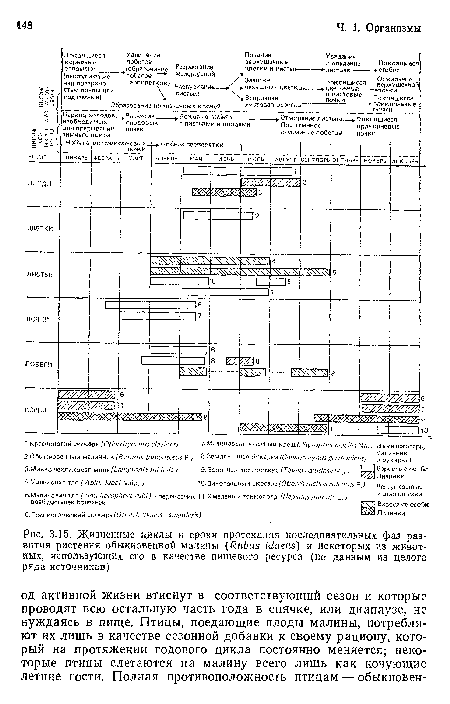 |
Состав различных частей растений и тел животных, используемых другими организмами в качестве пищевых ресурсов (по данным из целого ряда источников)
| Состав различных частей растений и тел животных, используемых другими организмами в качестве пищевых ресурсов (по данным из целого ряда источников) | 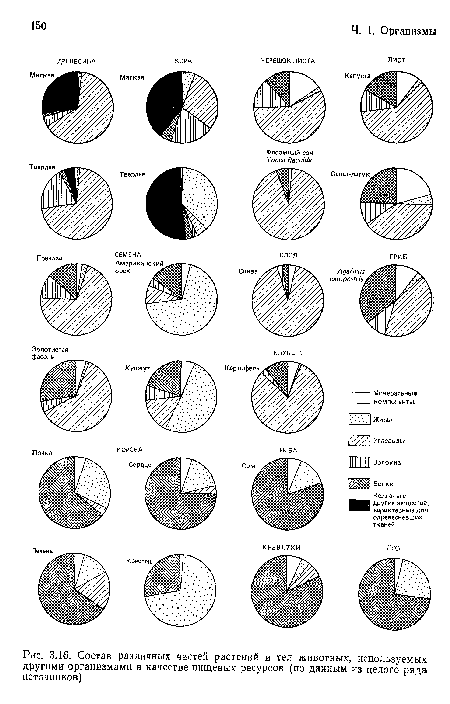 |
Голова кормящейся тли. Виден хоботок (стилет), проникающий во флоэму листа
| Голова кормящейся тли. Виден хоботок (стилет), проникающий во флоэму листа | 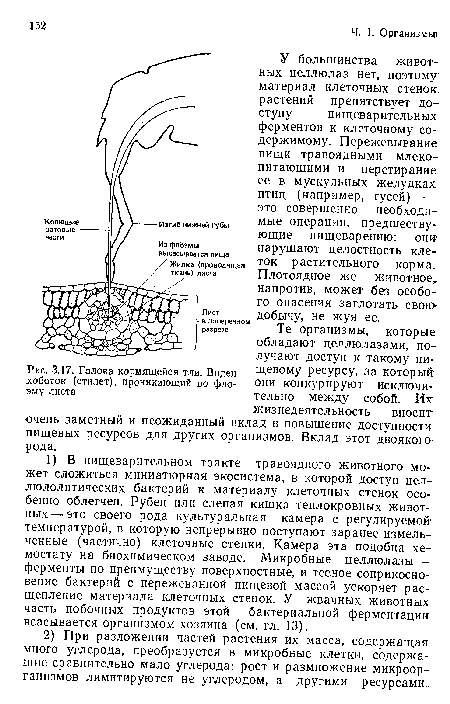 |
Коробочки маков (Рараюег эрр.) различных видов. У диких маков с гладкими коробочками поры на коробочках большие, и семена из них высыпаются быстро. Коробочки, защищенные покрывающими их колючими шипиками, принадлежат тем макам, у которых поры на коробочках мелкие; семена из таких пор высылаются медленно.
| Коробочки маков (Рараюег эрр.) различных видов. У диких маков с гладкими коробочками поры на коробочках большие, и семена из них высыпаются быстро. Коробочки, защищенные покрывающими их колючими шипиками, принадлежат тем макам, у которых поры на коробочках мелкие; семена из таких пор высылаются медленно. |  |
Среднее число самок весенней капустной мухи (Delia brassicae), ориентирующихся против ветра в направлении капустной грядки, в зависимости от расстояния, отделяющего их от ближайших растений капусты. (Hawkes, 1974.)
| Среднее число самок весенней капустной мухи (Delia brassicae), ориентирующихся против ветра в направлении капустной грядки, в зависимости от расстояния, отделяющего их от ближайших растений капусты. (Hawkes, 1974.) |  |
Изолинии скоростей роста, зависящих от степени обеспеченности ресурсами. Каждая из этих изолиний соединяет точки, соответствующие таким количествам двух ресурсов (#1 и #2) в окружающей среде, которые понадобились бы для того, чтобы популяция росла с данной скоростью (для каждой изолинии— своей). С ростом обилия ресурсов скорость роста популяции увеличивается, и поэтому чем выше скорость роста, тем дальше от начала координат расположена соответствующая ей изолиния
| Изолинии скоростей роста, зависящих от степени обеспеченности ресурсами. Каждая из этих изолиний соединяет точки, соответствующие таким количествам двух ресурсов (#1 и #2) в окружающей среде, которые понадобились бы для того, чтобы популяция росла с данной скоростью (для каждой изолинии— своей). С ростом обилия ресурсов скорость роста популяции увеличивается, и поэтому чем выше скорость роста, тем дальше от начала координат расположена соответствующая ей изолиния | 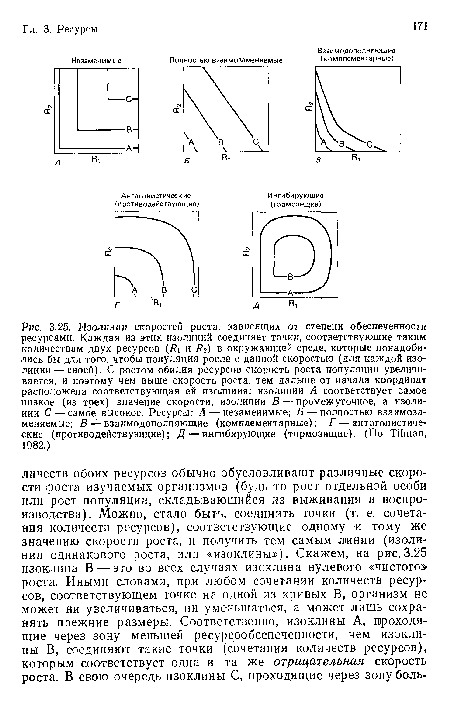 |
Изменчивость особей
| Изменчивость особей |  |
Различные модулярные организмы (слева — растения, справа — животные). А. Модулярные организмы, по мере роста распадающиеся на части
| Различные модулярные организмы (слева — растения, справа — животные). А. Модулярные организмы, по мере роста распадающиеся на части |  |
Возрастной состав клонов песчаной осоки Carex arenaria, растущих на песчаных дюнах Северного Уэльса (клоны состоят из разновозрастных побегов). Внесение минеральных удобрений приводит к тому, что этот возрастной состав изменяется
| Возрастной состав клонов песчаной осоки Carex arenaria, растущих на песчаных дюнах Северного Уэльса (клоны состоят из разновозрастных побегов). Внесение минеральных удобрений приводит к тому, что этот возрастной состав изменяется | 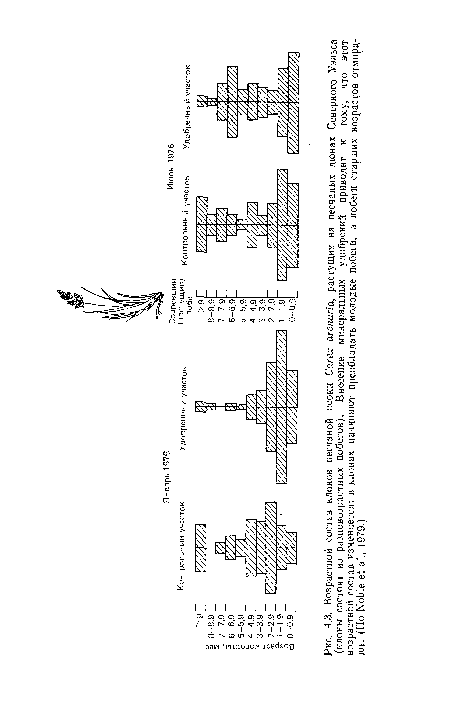 |
Изменения численности популяций. А. Овцы на о. Тасмания
| Изменения численности популяций. А. Овцы на о. Тасмания | 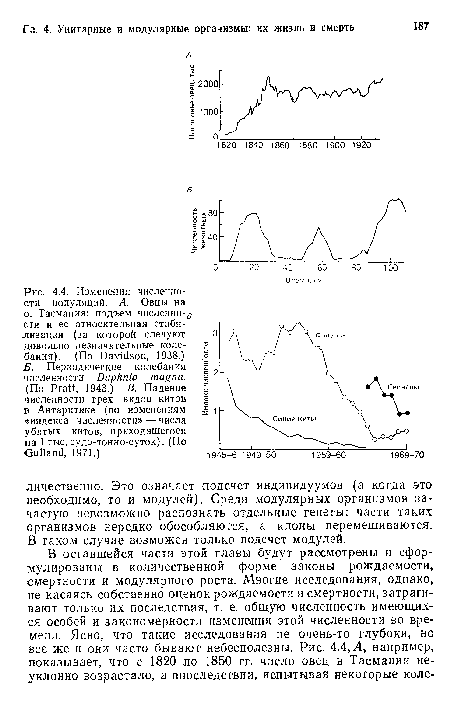 |
Различные типы жизненных циклов. Длина полосы в каждом случае отражает продолжительность жизни особи, а высота — размер тела. Толщина закрашенного фрагмента соответствует доле наличных ресурсов, направляемой на размножение. Организмы с однократным размножением уделяют ему большую часть ресурсов (во время размножения), но вскоре после этого отмирают. Организмы с многократным размножением во время очередной репродуктив-
| Различные типы жизненных циклов. Длина полосы в каждом случае отражает продолжительность жизни особи, а высота — размер тела. Толщина закрашенного фрагмента соответствует доле наличных ресурсов, направляемой на размножение. Организмы с однократным размножением уделяют ему большую часть ресурсов (во время размножения), но вскоре после этого отмирают. Организмы с многократным размножением во время очередной репродуктив- | 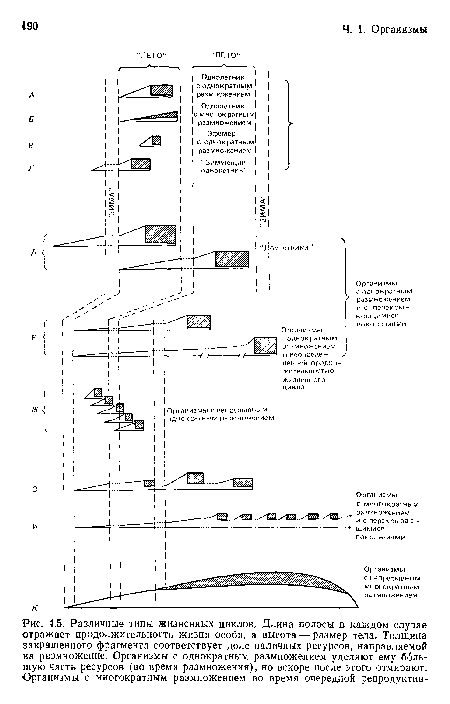 |
Между относительной численностью вегетирующих растений (гистограммы) и хранящихся в почве семян (вертикальные отрезки с горизонтальными полосками на концах) существуют значительные различия. Диаграммы отражают это несоответствие на примере наиболее многочисленных растений, растущих в листопадном лесу (Л) и на пашие (Б). (По Thompson, Grime, 1979.)
| Между относительной численностью вегетирующих растений (гистограммы) и хранящихся в почве семян (вертикальные отрезки с горизонтальными полосками на концах) существуют значительные различия. Диаграммы отражают это несоответствие на примере наиболее многочисленных растений, растущих в листопадном лесу (Л) и на пашие (Б). (По Thompson, Grime, 1979.) | 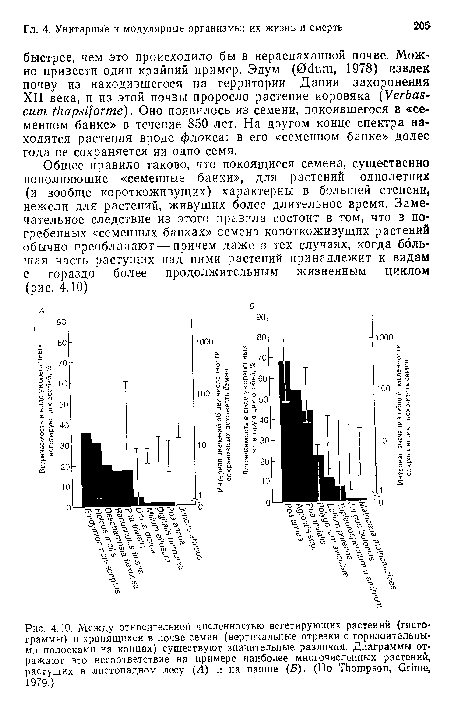 |
Две кривые выживания самок благородного оленя на острове Рам. Как объясняется в тексте, одна из этих кривых построена по когортной таблице выживания, соответствующей поколению 1957 г. рождения, и потому относится к периоду после 1957 г.; вторая же кривая построена по статической таблице выживания популяции, существовавшей в 1957 г., а потому относится к периоду до 1957 г. (по Lowe, 1969.) (На рисунке в обозначении кривых допущена ошибка
| Две кривые выживания самок благородного оленя на острове Рам. Как объясняется в тексте, одна из этих кривых построена по когортной таблице выживания, соответствующей поколению 1957 г. рождения, и потому относится к периоду после 1957 г.; вторая же кривая построена по статической таблице выживания популяции, существовавшей в 1957 г., а потому относится к периоду до 1957 г. (по Lowe, 1969.) (На рисунке в обозначении кривых допущена ошибка |  |
Кривая выживания когорты особей Acacia burkittii в Южной Австралии, полученная в результате регулярных подсчетов на одних и тех же участках (подсчитывались акации, взошедшие из семян в 1929 г.). (По Crisp, Lange, 1976.)
| Кривая выживания когорты особей Acacia burkittii в Южной Австралии, полученная в результате регулярных подсчетов на одних и тех же участках (подсчитывались акации, взошедшие из семян в 1929 г.). (По Crisp, Lange, 1976.) |  |
Возрастной состав популяций Acacia burkittii в двух местностях в Южной Австралии. (По Crisp, Lange, 1976.)
| Возрастной состав популяций Acacia burkittii в двух местностях в Южной Австралии. (По Crisp, Lange, 1976.) |  |
Повторяемость «кормных» лет. Колебания урожая шишек
| Повторяемость «кормных» лет. Колебания урожая шишек | 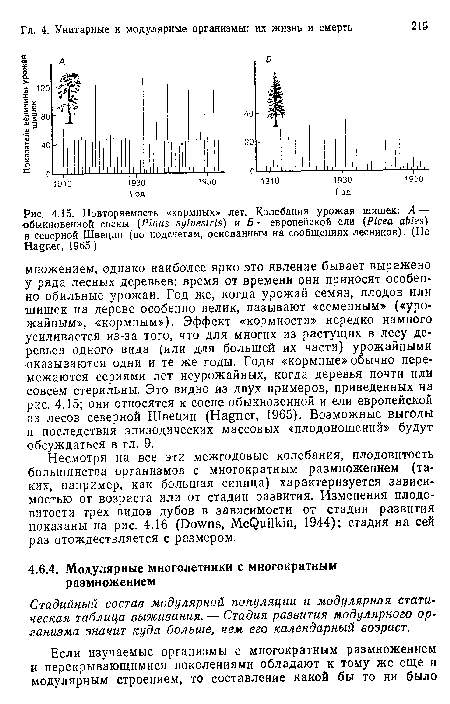 |
Зависимость плодовитости дубов трех видов от размеров деревьев; ДВГ — диаметр ствола на высоте груди. Кривые (отсчет ведется справа)
| Зависимость плодовитости дубов трех видов от размеров деревьев; ДВГ — диаметр ствола на высоте груди. Кривые (отсчет ведется справа) | 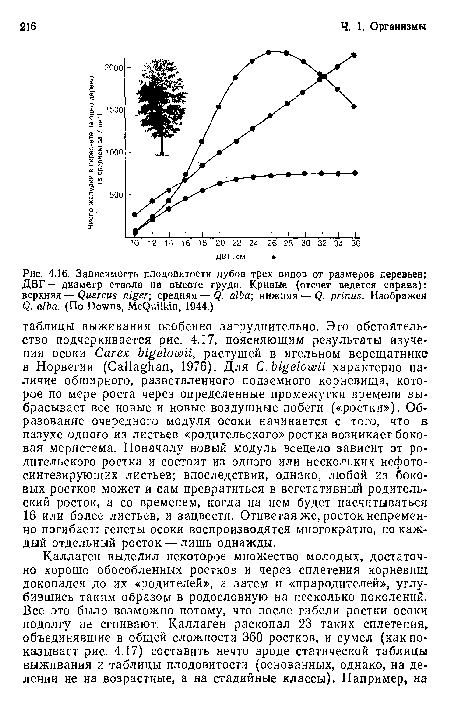 |
Связь между объемом побегов креозотового куста Larrea tri-dentata из долины Сан-Симон (Аризона) и его возрастом (возраст определялся по числу годовых колец на спиле). Креозотовый куст — модулярный организм, и поэтому оценить объем его побегов по возрасту очень трудно. (По Chew, Chew, 1965.)
| Связь между объемом побегов креозотового куста Larrea tri-dentata из долины Сан-Симон (Аризона) и его возрастом (возраст определялся по числу годовых колец на спиле). Креозотовый куст — модулярный организм, и поэтому оценить объем его побегов по возрасту очень трудно. (По Chew, Chew, 1965.) | 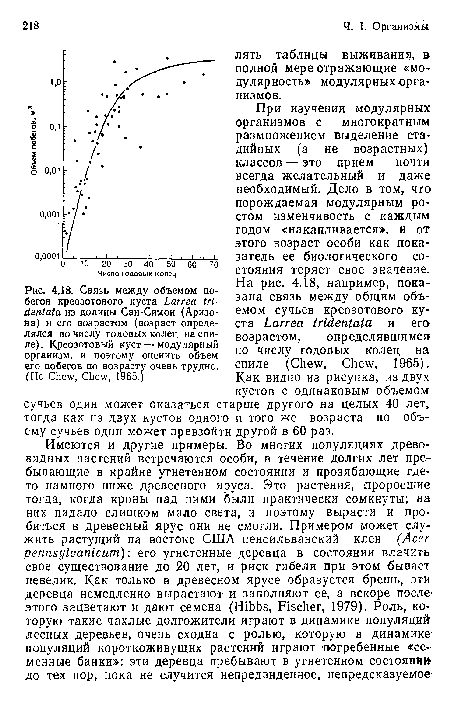 |
Общая численность и возрастной состав населения развивающихся и развитых стран (соответственно заштрихованные и незаштрихованные части диаграмм слева н справа). Верхняя диаграмма соответствует состоянию на 1980 год, а нижняя — прогнозу ООН на 2000 год. (По May, 1980.)
| Общая численность и возрастной состав населения развивающихся и развитых стран (соответственно заштрихованные и незаштрихованные части диаграмм слева н справа). Верхняя диаграмма соответствует состоянию на 1980 год, а нижняя — прогнозу ООН на 2000 год. (По May, 1980.) | 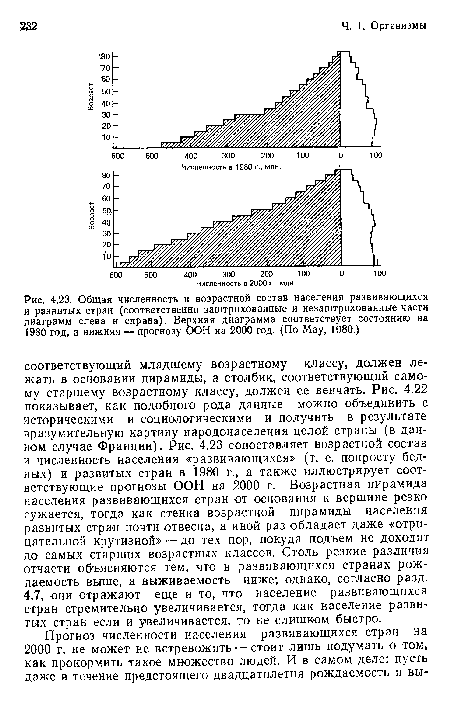 |
Три основных типа размещения организмов в пространстве (вверху) и соответствующие им распределения частот квадратных проб, содержащих то или иное число организмов (внизу). Эти распределения были бы получены при выборочном обследовании популяций с соответствующими типами размещения. А. Случайное размещение. Б. Регулярное размещение. В. Групповое размещение. На графиках (Б) и (В) для сравнения штриховой линией вычерчен также график (Л), характеризующий частотное распределение, соответствующее случайному размещению. При регулярном размещении число организмов в большей части квадратных проб примерно одно и то же. При групповом размещении обычно бывает довольно много квадратных проб с очень небольшим числом особей, а также довольно много таких проб, число особей в которых очень велико.
| Три основных типа размещения организмов в пространстве (вверху) и соответствующие им распределения частот квадратных проб, содержащих то или иное число организмов (внизу). Эти распределения были бы получены при выборочном обследовании популяций с соответствующими типами размещения. А. Случайное размещение. Б. Регулярное размещение. В. Групповое размещение. На графиках (Б) и (В) для сравнения штриховой линией вычерчен также график (Л), характеризующий частотное распределение, соответствующее случайному размещению. При регулярном размещении число организмов в большей части квадратных проб примерно одно и то же. При групповом размещении обычно бывает довольно много квадратных проб с очень небольшим числом особей, а также довольно много таких проб, число особей в которых очень велико. | 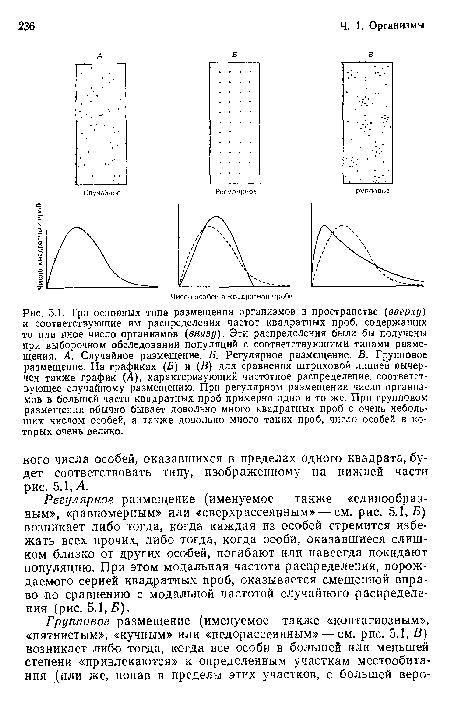 |
Закономерности миграций в связи с жизненными циклами
| Закономерности миграций в связи с жизненными циклами |  |
Миграционные пути полярной крачки (Sterna paradisaea). (Из> Kullenberg, 1946.)
| Миграционные пути полярной крачки (Sterna paradisaea). (Из> Kullenberg, 1946.) |  |
Рассеяние семян Eucalyptus regnans с деревьев высотой около 75 м, растущих на опушке густого леса (Л), и с одиноко растущего дерева (Б). В. Связь между относительной плотностью семейного покрова и относительной удаленностью от источника семян для ряда растений
| Рассеяние семян Eucalyptus regnans с деревьев высотой около 75 м, растущих на опушке густого леса (Л), и с одиноко растущего дерева (Б). В. Связь между относительной плотностью семейного покрова и относительной удаленностью от источника семян для ряда растений |  |
Диморфизм животных и растений по расселяемости. Крылатые и бескрылые самки
| Диморфизм животных и растений по расселяемости. Крылатые и бескрылые самки |  |
А. Обобщенная диаграмма, поясняющая зависимость успешности скрещивания (числа потомков и их жизнеспособности) от степени генетического сходства между брачными партнерами. Б. Результаты скрещивания дельфиниумов (Delphinium nelsoni) путем целенаправленного ручного искусственного опыления. Они зависят от расстояния, разделявшего на местности родительские растения. (Из Price, Waser, 1979). Черными и белыми кружками обозначены результаты двух различных опытов.
| А. Обобщенная диаграмма, поясняющая зависимость успешности скрещивания (числа потомков и их жизнеспособности) от степени генетического сходства между брачными партнерами. Б. Результаты скрещивания дельфиниумов (Delphinium nelsoni) путем целенаправленного ручного искусственного опыления. Они зависят от расстояния, разделявшего на местности родительские растения. (Из Price, Waser, 1979). Черными и белыми кружками обозначены результаты двух различных опытов. | 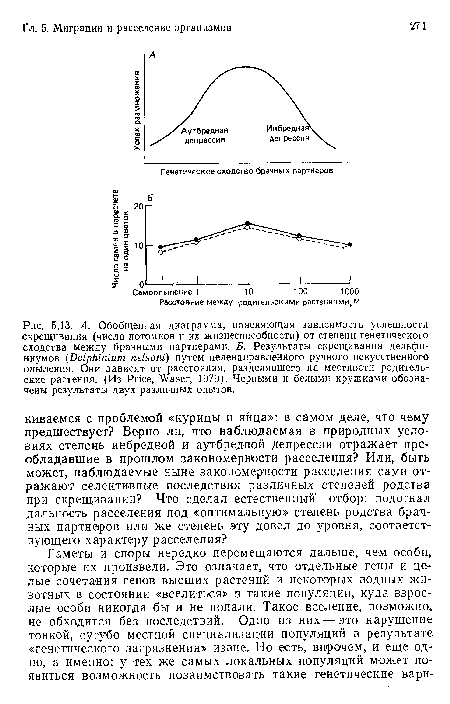 |
Зависимая от плотности смертность мальков форели. При высокой плотности популяции увеличение смертности точно компенсирует возрастание плотности, результатом чего является постоянная численность выживающих особей. (Из Hassel, 1976, по Le Cren, 1973.)
| Зависимая от плотности смертность мальков форели. При высокой плотности популяции увеличение смертности точно компенсирует возрастание плотности, результатом чего является постоянная численность выживающих особей. (Из Hassel, 1976, по Le Cren, 1973.) |  |
Зависимые от плотности рождаемость и смертность обусловливают регуляцию численности популяции. Если оба (Л) или один из параметров (Б и В) зависят от плотности, то кривые пересекаются. Плотность, при которой происходит пересечение, называется предельной плотностью насыщения (К). При плотности ниже К популяция растет, при плотности выше К — уменьшается; при плотности, равной К, наблюдается устойчивое равновесие. На рисунке дана лишь самая общая схема
| Зависимые от плотности рождаемость и смертность обусловливают регуляцию численности популяции. Если оба (Л) или один из параметров (Б и В) зависят от плотности, то кривые пересекаются. Плотность, при которой происходит пересечение, называется предельной плотностью насыщения (К). При плотности ниже К популяция растет, при плотности выше К — уменьшается; при плотности, равной К, наблюдается устойчивое равновесие. На рисунке дана лишь самая общая схема | 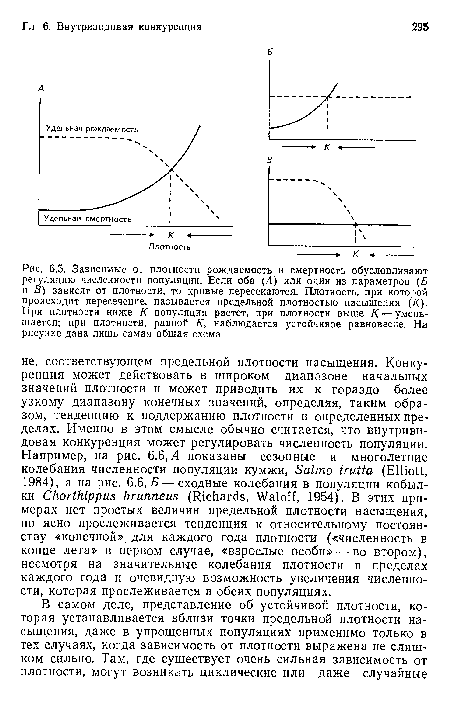 |
Некоторые общие аспекты внутривидовой конкуренции. А. Зависимые от плотности изменения числа погибших и родившихся особей в популяции
| Некоторые общие аспекты внутривидовой конкуренции. А. Зависимые от плотности изменения числа погибших и родившихся особей в популяции | 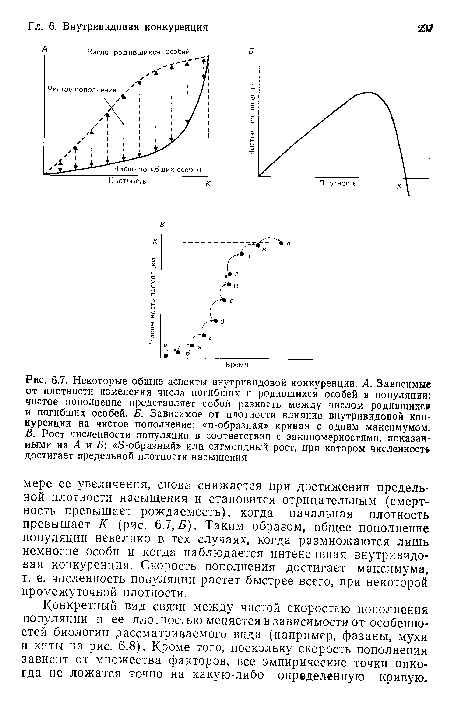 |
Внутривидовая конкуренция и рост в популяциях блюдечка Patella cochlear. А. С ростом плотности размеры особей уменьшаются, что приводит к поддержанию общей биомассы популяции на постоянном уровне. Б. В популяциях с высокой плотностью много мелких и мало крупных особей; при низкой плотности популяция состоит из большого числа крупных и небольшого числа мелких особей. (По Branch, 1975.)
| Внутривидовая конкуренция и рост в популяциях блюдечка Patella cochlear. А. С ростом плотности размеры особей уменьшаются, что приводит к поддержанию общей биомассы популяции на постоянном уровне. Б. В популяциях с высокой плотностью много мелких и мало крупных особей; при низкой плотности популяция состоит из большого числа крупных и небольшого числа мелких особей. (По Branch, 1975.) | 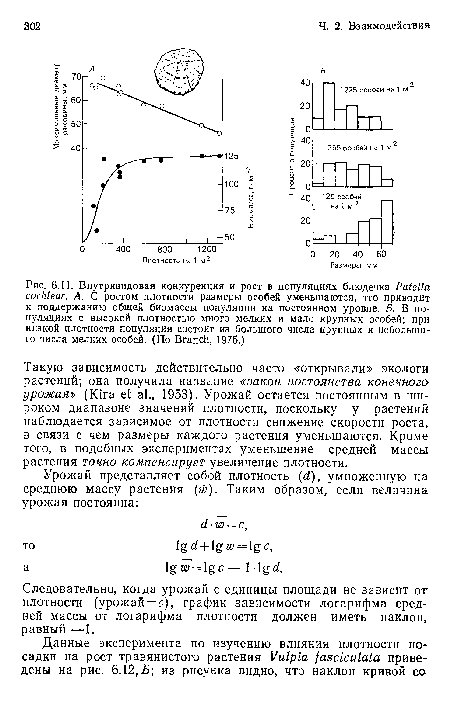 |
Конкуренция в посевах кукурузы (Zea mays) гораздо сильнее влияет на продукцию семян (масса початков), чем на рост побегов. (Harper, 1961.)
| Конкуренция в посевах кукурузы (Zea mays) гораздо сильнее влияет на продукцию семян (масса початков), чем на рост побегов. (Harper, 1961.) | 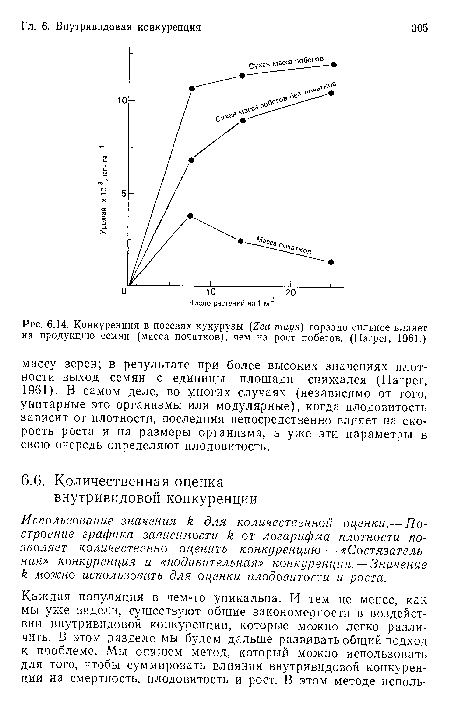 |
Математические модели роста численности популяции с дискретными поколениями
| Математические модели роста численности популяции с дискретными поколениями |  |
Простейшая прямолинейная зависимость, в соответствии с которой с возрастанием плотности (А^<) может увеличиваться величина, обратная показателю роста генерации ( Л^/Л^+1). Подробнее см. в тексте
| Простейшая прямолинейная зависимость, в соответствии с которой с возрастанием плотности (А^<) может увеличиваться величина, обратная показателю роста генерации ( Л^/Л^+1). Подробнее см. в тексте | 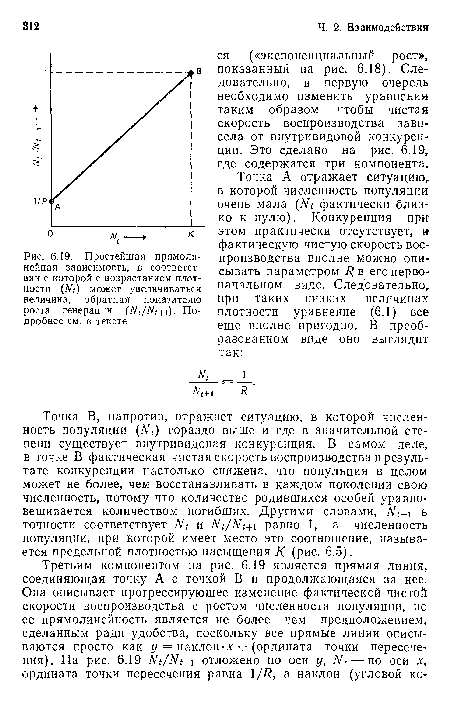 |
Внутривидовая конкуренция, выраженная уравнением 6.3. Независимо от начальной плотности или константы а=(к— )/К наклон линии, связывающей Л
| Внутривидовая конкуренция, выраженная уравнением 6.3. Независимо от начальной плотности или константы а=(к— )/К наклон линии, связывающей Л | 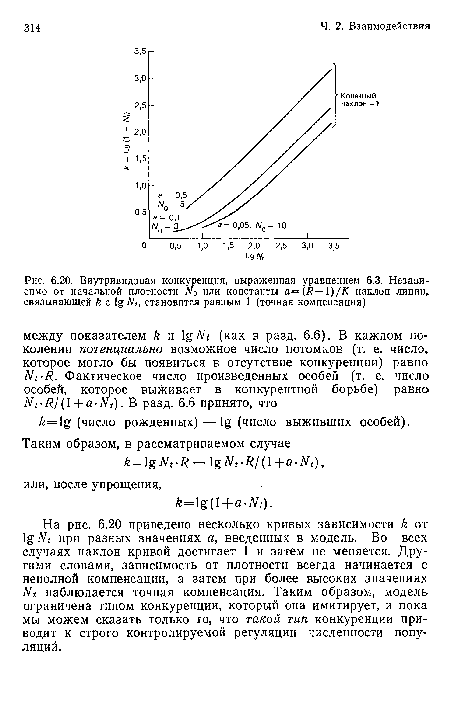 |
Внутривидовая конкуренция, выраженная уравнением 6.4. Конечный наклон определяется значением Ь в этом уравнении.
| Внутривидовая конкуренция, выраженная уравнением 6.4. Конечный наклон определяется значением Ь в этом уравнении. | 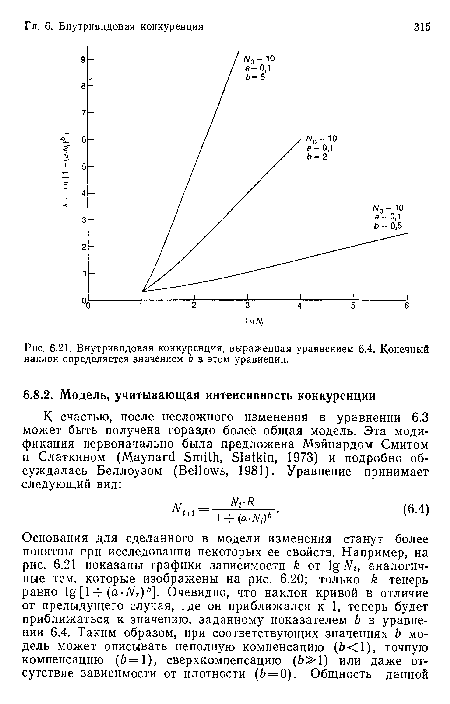 |
А. Различные варианты динамики численности популяции (показаны на рис. Б), полученные с помощью уравнения 6.4 при разном сочетании параметров Ь и R. Разделение устойчивых предельных циклов и случайных изменений является приблизительным. (По May, 1975а и Bellows, 1981.)
| А. Различные варианты динамики численности популяции (показаны на рис. Б), полученные с помощью уравнения 6.4 при разном сочетании параметров Ь и R. Разделение устойчивых предельных циклов и случайных изменений является приблизительным. (По May, 1975а и Bellows, 1981.) | 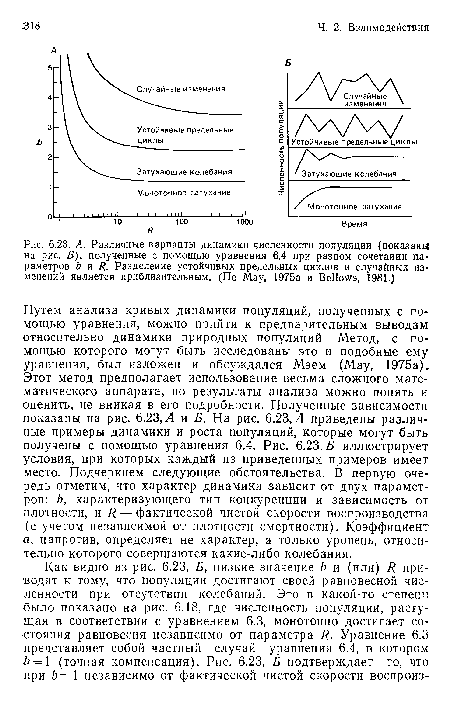 |
Экспоненциальное и Б-образное увеличение плотности (ЛО во времени в моделях с непрерывным размножением. Б-образный рост описывается логистическим уравнением
| Экспоненциальное и Б-образное увеличение плотности (ЛО во времени в моделях с непрерывным размножением. Б-образный рост описывается логистическим уравнением |  |
Простейшая прямолинейная зависимость, иллюстрирующая снижение удельной скорости роста (<1ЛГ/<М 1/ЛГ) в связи с увеличением плотности (ЛГ). В тексте приводится более подробное обсуждение
| Простейшая прямолинейная зависимость, иллюстрирующая снижение удельной скорости роста (<1ЛГ/<М 1/ЛГ) в связи с увеличением плотности (ЛГ). В тексте приводится более подробное обсуждение |  |
Конкуренция за пространство у многолетнего растения Anemone hepatica в лесах Швеции. Каждому экземпляру соответствует одна линия
| Конкуренция за пространство у многолетнего растения Anemone hepatica в лесах Швеции. Каждому экземпляру соответствует одна линия |  |
Возможные затраты, выигрыш и чистый выигрыш (выигрыш минус затраты) при защите индивидуального участка. При переходе к территориальности затраты резко увеличиваются и затем они увеличиваются со все более высокой скоростью по мере того, как возрастает сложность защиты индивидуального участка. Если на индивидуальном участке содержится все необходимое для организма, то выигрыш перестает расти. Если плотность ресурса (в данном случае нектар) высока, то это происходит при небольших размерах индивидуального участка, а если плотность ресурса низкая, то при больших размерах. Вследствие этого при более высокой плотности ресурса чистый выигрыш достигает максимальной величины на индивидуальном участке меньшего размера
| Возможные затраты, выигрыш и чистый выигрыш (выигрыш минус затраты) при защите индивидуального участка. При переходе к территориальности затраты резко увеличиваются и затем они увеличиваются со все более высокой скоростью по мере того, как возрастает сложность защиты индивидуального участка. Если на индивидуальном участке содержится все необходимое для организма, то выигрыш перестает расти. Если плотность ресурса (в данном случае нектар) высока, то это происходит при небольших размерах индивидуального участка, а если плотность ресурса низкая, то при больших размерах. Вследствие этого при более высокой плотности ресурса чистый выигрыш достигает максимальной величины на индивидуальном участке меньшего размера |  |
Явление самоизреживания среди различных видов трав и деревьев. Каждая линия представляет отдельный вид и в то же время обозначает диапазон значений, в котором производились наблюдения. Стрелками на отдельных линиях обозначено направление, в котором происходит самоизрежи-ваиие во времени. Графики построены по данным, приведенным на рис. 2.9, из работы Уайта (White, 1980), в которой имеются ссылки на источники и перечислены все растения (31 вид). Обратите внимание, что все линии имеют наклон, примерно равный — 3/г, а их параметры попадают в относительно узкий диапазон значений
| Явление самоизреживания среди различных видов трав и деревьев. Каждая линия представляет отдельный вид и в то же время обозначает диапазон значений, в котором производились наблюдения. Стрелками на отдельных линиях обозначено направление, в котором происходит самоизрежи-ваиие во времени. Графики построены по данным, приведенным на рис. 2.9, из работы Уайта (White, 1980), в которой имеются ссылки на источники и перечислены все растения (31 вид). Обратите внимание, что все линии имеют наклон, примерно равный — 3/г, а их параметры попадают в относительно узкий диапазон значений | 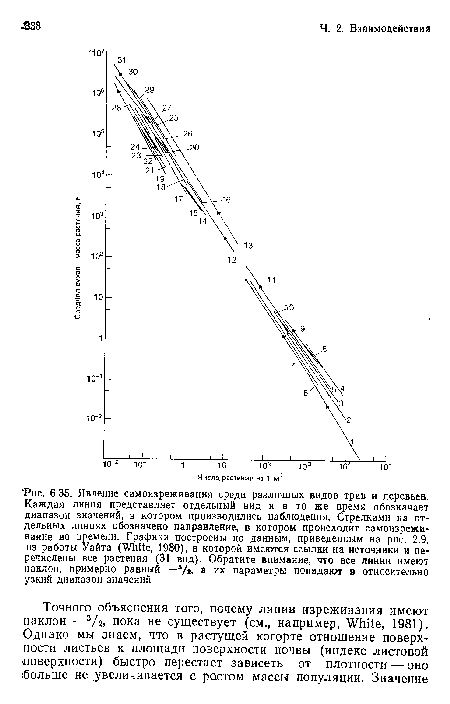 |
Изоклины, полученные с помощью уравнений конкуренции Лотки— Вольтерры. А. Изоклина для Л^
| Изоклины, полученные с помощью уравнений конкуренции Лотки— Вольтерры. А. Изоклина для Л^ |  |
А и Б иллюстрируют те случаи, в которых сильный конкурент неизменно побеждает более слабого. Эту ситуацию полезно рассмотреть с точки зрения теории экологической ниши (разд. 2.12 и 3.7). Вспомним, что в отсутствие конкурентов другого вида экологическая ниша представляет собой фундаментальную нишу (определенную сочетанием условий и ресурсов, позволяющим виду поддерживать жизнеспособную популяцию). Однако в присутствии конкурентов виду приходится довольствоваться реализованной нишей, конкретные свойства которой определяются конкурирующими видами. Это различие подчеркивает, что межвидовая конкуренция приводит к снижению плодовитости и жизнеспособности и что в фундаментальной экологической нише может быть такая часть, занимая которую вид в результате межвидовой конкуренции не в состоянии больше жить и успешно размножаться. Эта часть фундаментальной ниши вида отсутствует в его реализованной нише. Возвращаясь к рис. 7.8, Л и Б, мы, таким образом, можем сказать, что, конкурируя с более сильным видом, слабый конкурент утрачивает свою реализованную нишу. Обсуждавшиеся до этого примеры межвидовой конкуренции теперь могут быть рассмотрены в рамках понятия экологической ниши.
| А и Б иллюстрируют те случаи, в которых сильный конкурент неизменно побеждает более слабого. Эту ситуацию полезно рассмотреть с точки зрения теории экологической ниши (разд. 2.12 и 3.7). Вспомним, что в отсутствие конкурентов другого вида экологическая ниша представляет собой фундаментальную нишу (определенную сочетанием условий и ресурсов, позволяющим виду поддерживать жизнеспособную популяцию). Однако в присутствии конкурентов виду приходится довольствоваться реализованной нишей, конкретные свойства которой определяются конкурирующими видами. Это различие подчеркивает, что межвидовая конкуренция приводит к снижению плодовитости и жизнеспособности и что в фундаментальной экологической нише может быть такая часть, занимая которую вид в результате межвидовой конкуренции не в состоянии больше жить и успешно размножаться. Эта часть фундаментальной ниши вида отсутствует в его реализованной нише. Возвращаясь к рис. 7.8, Л и Б, мы, таким образом, можем сказать, что, конкурируя с более сильным видом, слабый конкурент утрачивает свою реализованную нишу. Обсуждавшиеся до этого примеры межвидовой конкуренции теперь могут быть рассмотрены в рамках понятия экологической ниши. |  |
Кривые использования ресурса для трех видов, сосуществующих в одномерном пространстве ресурса. d — расстояние между соседними максимумами; w — стандартное отклонение для кривых. А. Узкие ниши со слабым перекрыванием (d>w), т. е. межвидовая конкуренция сравнительно невелика. Б. Более широкие ниши с сильным перекрыванием (d<Lw), т. е. имеет место сравнительно интенсивная межвидовая конкуренция.
| Кривые использования ресурса для трех видов, сосуществующих в одномерном пространстве ресурса. d — расстояние между соседними максимумами; w — стандартное отклонение для кривых. А. Узкие ниши со слабым перекрыванием (d>w), т. е. межвидовая конкуренция сравнительно невелика. Б. Более широкие ниши с сильным перекрыванием (d<Lw), т. е. имеет место сравнительно интенсивная межвидовая конкуренция. |  |
Область благоприятных условий среды (обозначена величинами предельной плотности насыщения К и К% где К = Кз, при которой возможно существование равновесного сообщества из трех видов с разной степенью перекрывания ниш (d/w). (По May, 1973.)
| Область благоприятных условий среды (обозначена величинами предельной плотности насыщения К и К% где К = Кз, при которой возможно существование равновесного сообщества из трех видов с разной степенью перекрывания ниш (d/w). (По May, 1973.) | 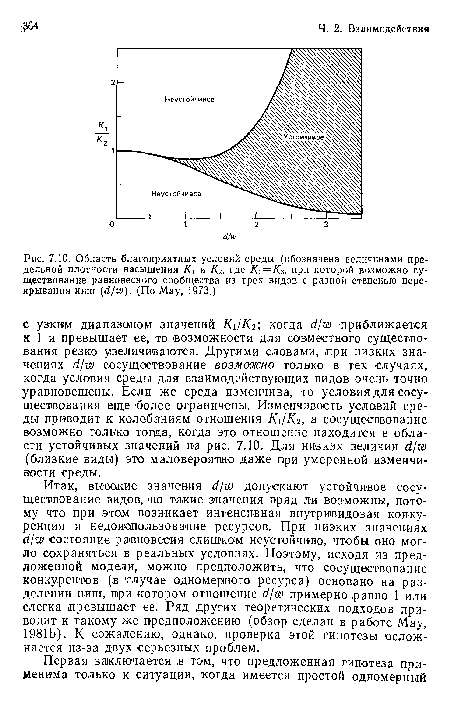 |
Формальные схемы взаимодействий
| Формальные схемы взаимодействий | 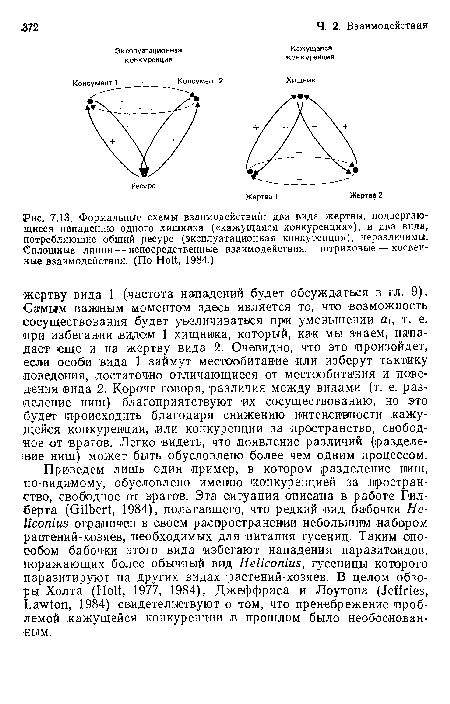 |
Эксперименты по конкуренции при возрастающей общей плотности посева
| Эксперименты по конкуренции при возрастающей общей плотности посева | 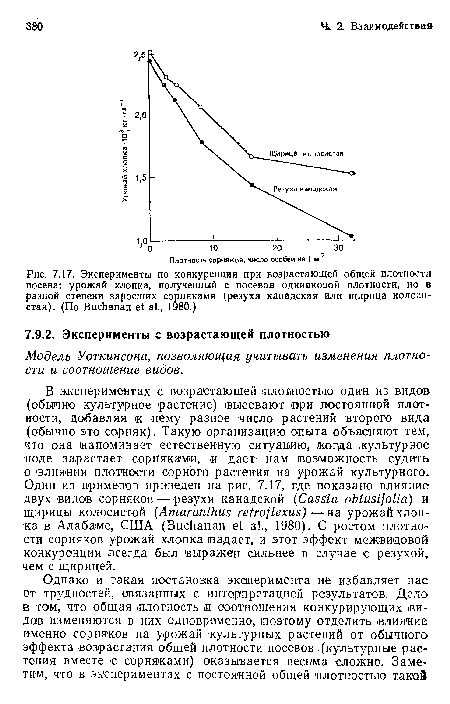 |
Зависимость числа семян на одном растении от плотности плодоносящих растений Agrostemma в монокультуре (кружки) и в посевах с пшеницей (квадраты). Для смешанных посевов уравнение кривой выглядит так
| Зависимость числа семян на одном растении от плотности плодоносящих растений Agrostemma в монокультуре (кружки) и в посевах с пшеницей (квадраты). Для смешанных посевов уравнение кривой выглядит так | 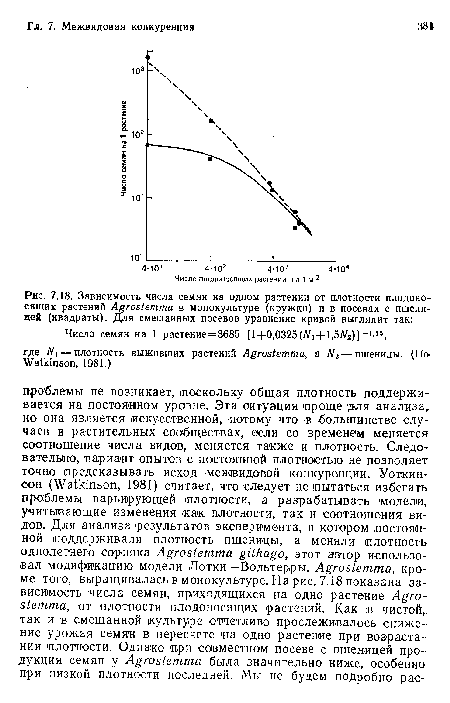 |
Смещение признаков
| Смещение признаков | 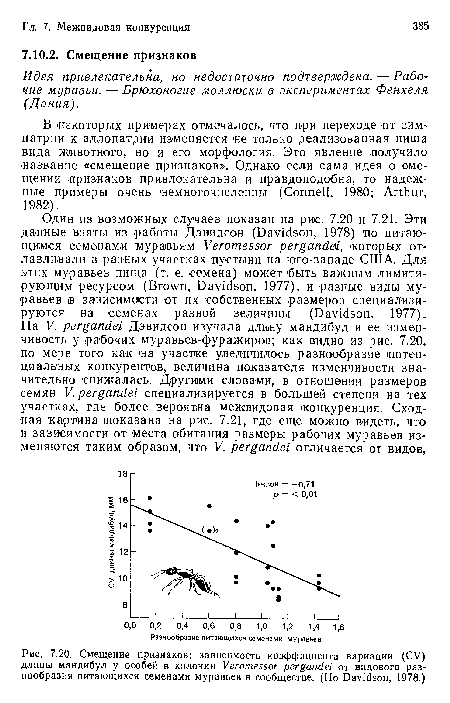 |
Смещение признаков
| Смещение признаков | 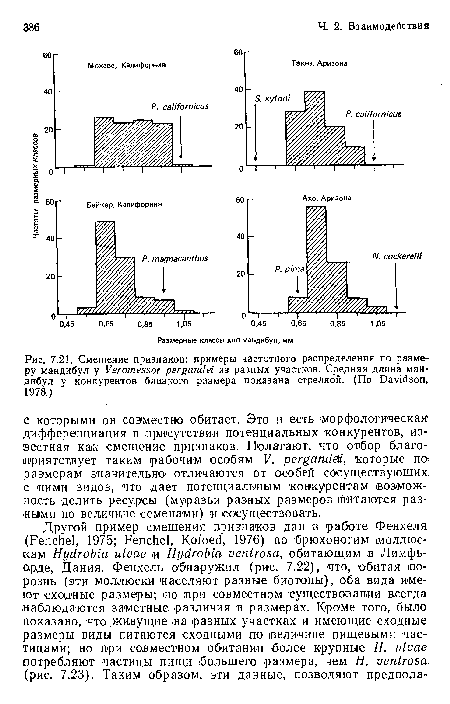 |
Смещение признаков
| Смещение признаков | 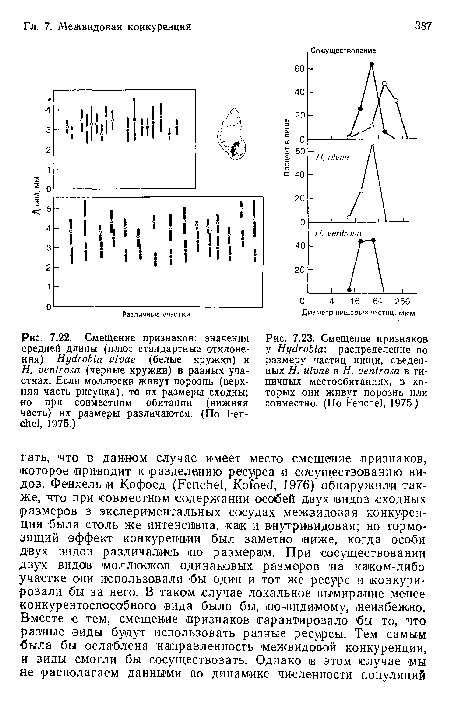 |
Конкурентное исключение
| Конкурентное исключение | 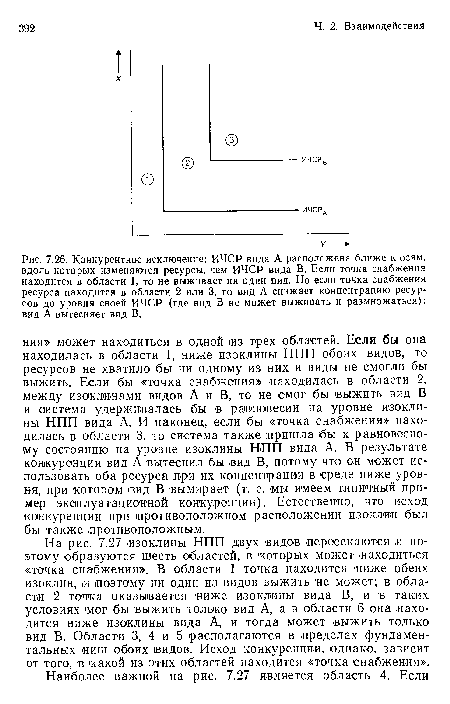 |
Возможности сосуществования двух конкурентов, ограниченных двумя необходимыми ресурсами. ИЧСР видов А и В перекрываются, что приводит к появлению шести различных областей. Если точка снабжения находится в области 1, то не выживает ни одии из видов; если точки расположены в областях 2 и 3, то вид А вытесняет вид В; а если в областях 5 и 6, то вид В вытесняет вид А. При размещении точек снабжения в области 4, которая ограничена двумя штриховыми линиями, виды сосуществуют. Более подробные объяснения см. в тексте.
| Возможности сосуществования двух конкурентов, ограниченных двумя необходимыми ресурсами. ИЧСР видов А и В перекрываются, что приводит к появлению шести различных областей. Если точка снабжения находится в области 1, то не выживает ни одии из видов; если точки расположены в областях 2 и 3, то вид А вытесняет вид В; а если в областях 5 и 6, то вид В вытесняет вид А. При размещении точек снабжения в области 4, которая ограничена двумя штриховыми линиями, виды сосуществуют. Более подробные объяснения см. в тексте. | 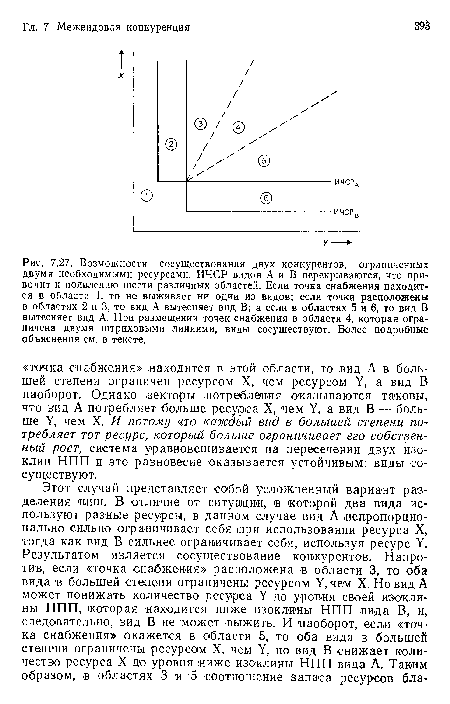 |
Урожайность семян
| Урожайность семян | 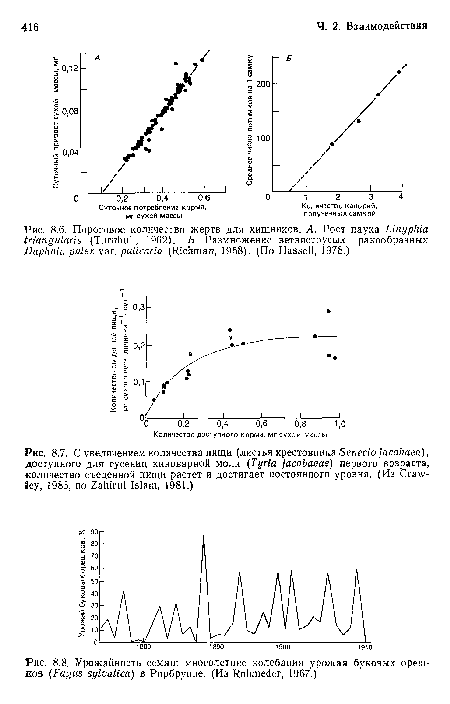 |
Переключение. А. Отсутствие переключения
| Переключение. А. Отсутствие переключения | 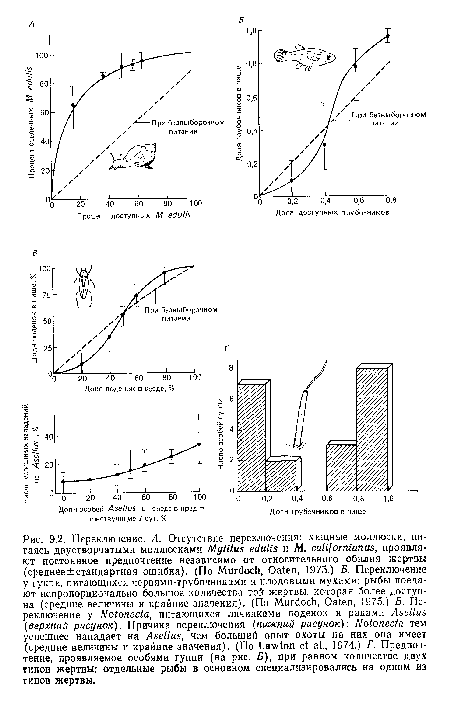 |
Сезонные изменения
| Сезонные изменения | 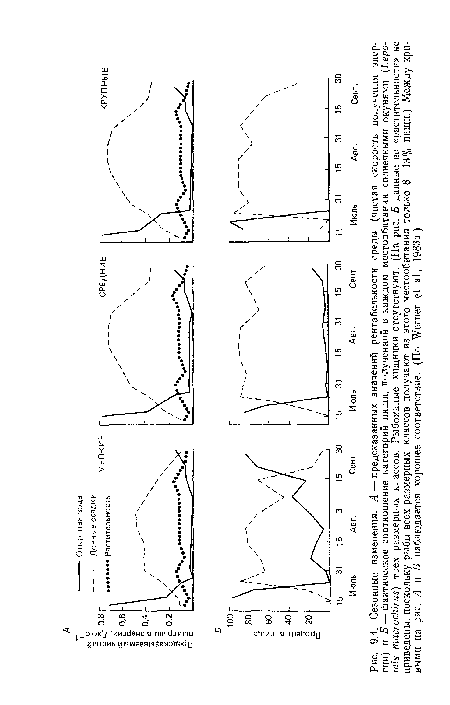 |
Взаимная интерференция
| Взаимная интерференция |  |
Взаимная интерференция
| Взаимная интерференция |  |
Колюшки (небольшая рыба) размещаются в соответствии с моделью идеального свободного распределения. В каждом случае использованы шесть рыб, которые могли кормиться в «более рентабельном» и «менее рентабельном» концах аквариума. Точки и вертикальные линии — средние величины и стандартные отклонения по 8 повторностям (Л) и 11 повторностям (Б). А. Число рыб, которые кормятся в менее рентабельном конце при соотношении показателей рентабельности 5
| Колюшки (небольшая рыба) размещаются в соответствии с моделью идеального свободного распределения. В каждом случае использованы шесть рыб, которые могли кормиться в «более рентабельном» и «менее рентабельном» концах аквариума. Точки и вертикальные линии — средние величины и стандартные отклонения по 8 повторностям (Л) и 11 повторностям (Б). А. Число рыб, которые кормятся в менее рентабельном конце при соотношении показателей рентабельности 5 |  |
А. Попав в кормовое пятно, личинка Р1еЫгоспепиа сопзрегва пятого возраста, которая встречает и съедает хирономиду, тотчас же прекращает поиск и начинает постройку ловчей сети. Хищники, которые не встретили жертву в течение 30 мин эксперимента, ведут более активный поиск, поэтому с большей вероятностью покинут пятно.
| А. Попав в кормовое пятно, личинка Р1еЫгоспепиа сопзрегва пятого возраста, которая встречает и съедает хирономиду, тотчас же прекращает поиск и начинает постройку ловчей сети. Хищники, которые не встретили жертву в течение 30 мин эксперимента, ведут более активный поиск, поэтому с большей вероятностью покинут пятно. | 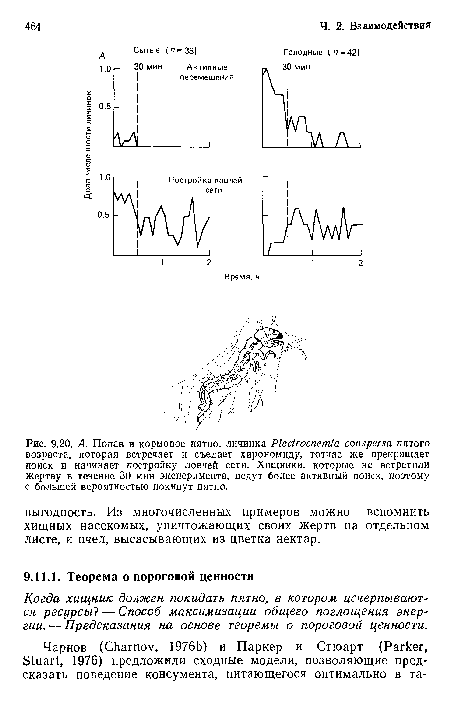 |
Теорема о пороговой ценности. А. Когда хищник попадает в кормовое пятно, скорость получения им энергии вначале высока (особенно в высокопродуктивных пятнах или там, где высока эффективность добывания пищи хищником), но со временем, по мере того как пятно истощается, эта скорость падает. Кривая совокупного получения энергии достигает асимптоты. Б. Выбор возможностей для хнщннка. Сплошная кривая представляет собой совокупную энергию, полученную в пятне средней продуктивности, & — среднее время перемещения от одного пятна к другому. Скорость получения энергии (которая должна быть максимизирована) — это количество полученной энергии, отнесенное к общему времени, т. е. наклон отрезка, проведеииого из начала координат до пересечения с кривой. Как кратковременное (наклои= =£’кр/(</+5кр), так и длительное [наклон=£,дл(^-Ь5дл)1 пребывания хищника в кормовом пятне приводят к более низкой скорости получения энергии (меиь-
| Теорема о пороговой ценности. А. Когда хищник попадает в кормовое пятно, скорость получения им энергии вначале высока (особенно в высокопродуктивных пятнах или там, где высока эффективность добывания пищи хищником), но со временем, по мере того как пятно истощается, эта скорость падает. Кривая совокупного получения энергии достигает асимптоты. Б. Выбор возможностей для хнщннка. Сплошная кривая представляет собой совокупную энергию, полученную в пятне средней продуктивности, & — среднее время перемещения от одного пятна к другому. Скорость получения энергии (которая должна быть максимизирована) — это количество полученной энергии, отнесенное к общему времени, т. е. наклон отрезка, проведеииого из начала координат до пересечения с кривой. Как кратковременное (наклои= =£’кр/(</+5кр), так и длительное [наклон=£,дл(^-Ь5дл)1 пребывания хищника в кормовом пятне приводят к более низкой скорости получения энергии (меиь- | 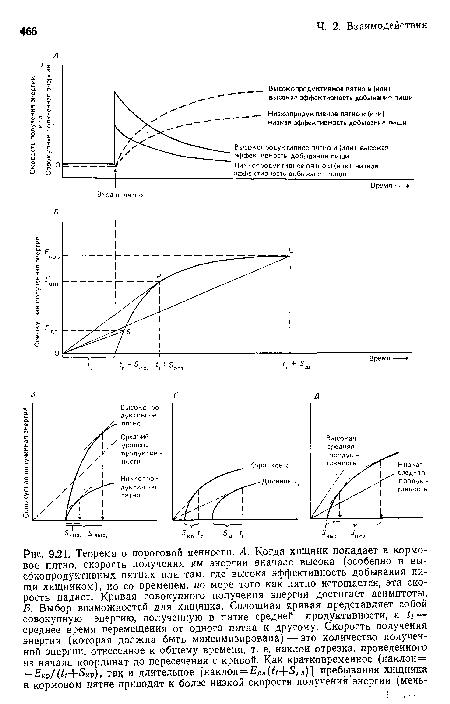 |
А. Экспериментальное «дерево» с тремя кормовыми пятнами для больших синиц. Б. Теоретическая зависимость оптимального времени пребывания в пятне от времени перехода между пятнами (штриховая линия) и наблюдавшиеся средние значения (¿стандартная ошибка) для шести птиц, каждая из которых испытывалась при двух условиях среды. В. Те же экспериментальные данные и теоретическая кривая, при построении которой учтены затраты энергии на перемещения между пятнами. (Из Krebs, 1978, по Cowie, 1977.)
| А. Экспериментальное «дерево» с тремя кормовыми пятнами для больших синиц. Б. Теоретическая зависимость оптимального времени пребывания в пятне от времени перехода между пятнами (штриховая линия) и наблюдавшиеся средние значения (¿стандартная ошибка) для шести птиц, каждая из которых испытывалась при двух условиях среды. В. Те же экспериментальные данные и теоретическая кривая, при построении которой учтены затраты энергии на перемещения между пятнами. (Из Krebs, 1978, по Cowie, 1977.) | 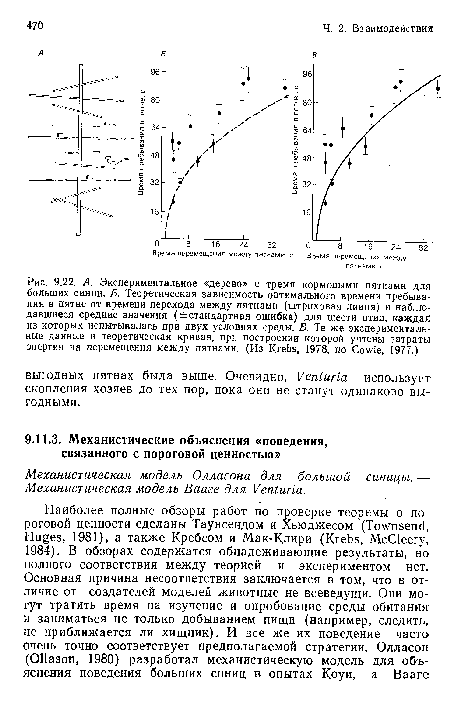 |
Данные Ссуиче (1977) по большой синице (см. рис. 9.22) в сравнении с прогнозом механистической модели памяти. (ОПавоп, 1980.)
| Данные Ссуиче (1977) по большой синице (см. рис. 9.22) в сравнении с прогнозом механистической модели памяти. (ОПавоп, 1980.) | 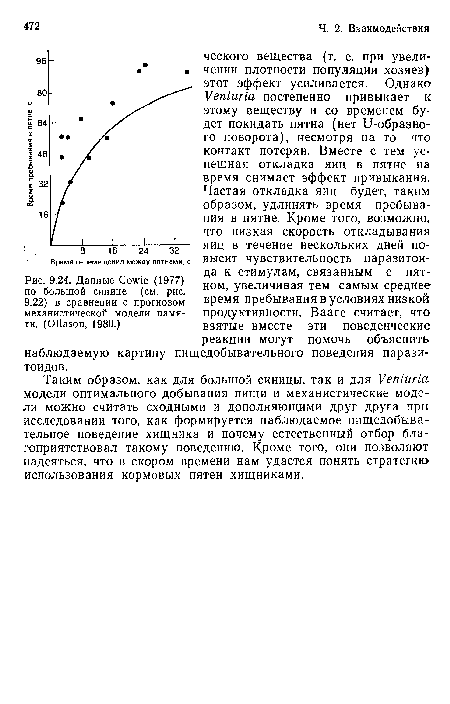 |
Модель Лотки—Вольтерры для системы хищник—жертва. А. Изоклина для популяции жертвы. При низкой плотности хищника (С) численность жертвы (М) возрастает, а при более высокой — снижается. Б. Изоклина для популяции хищника. При высокой плотности жертвы численность популяции хищника растет, а при низкой — снижается. В. При объединении изоклин в системе хищник—жертва возникают неограниченные взаимосвязанные колебания численности, подобные тем, которые показаны на рис. Г. Однако, как видно на рис. Д, для этих колебаний характерна нейтральная стабильность
| Модель Лотки—Вольтерры для системы хищник—жертва. А. Изоклина для популяции жертвы. При низкой плотности хищника (С) численность жертвы (М) возрастает, а при более высокой — снижается. Б. Изоклина для популяции хищника. При высокой плотности жертвы численность популяции хищника растет, а при низкой — снижается. В. При объединении изоклин в системе хищник—жертва возникают неограниченные взаимосвязанные колебания численности, подобные тем, которые показаны на рис. Г. Однако, как видно на рис. Д, для этих колебаний характерна нейтральная стабильность | 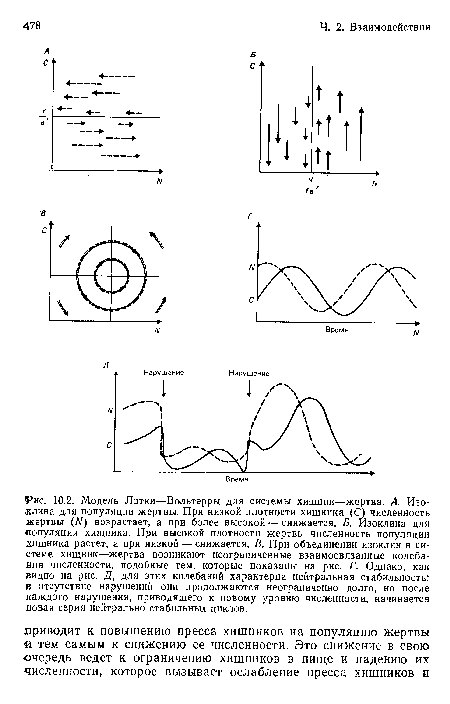 |
Зависимость от плотности с запаздыванием. А. Модель взаимодействия паразитоид—хозяин, исследованная на протяжении 50 поколений (Hassell, 1985)
| Зависимость от плотности с запаздыванием. А. Модель взаимодействия паразитоид—хозяин, исследованная на протяжении 50 поколений (Hassell, 1985) | 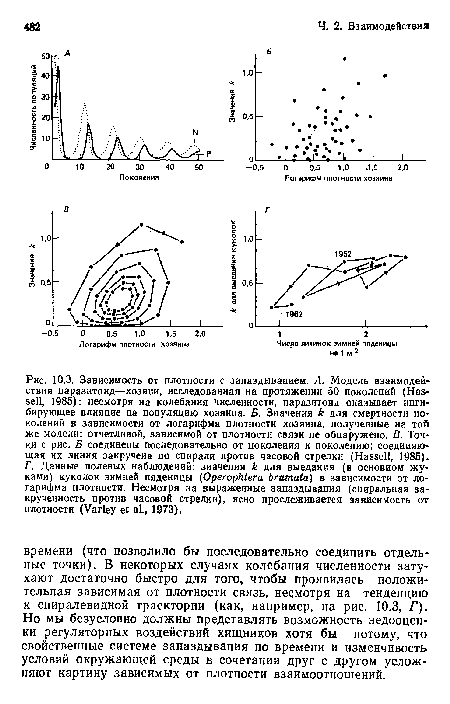 |
Взаимосвязанные колебания численности в лабораторной системе хищник—жертва
| Взаимосвязанные колебания численности в лабораторной системе хищник—жертва |  |
Колебания относительных величин биомассы основных компонентов «10-летнего природного цикла» в провинции Альберта, Канада. Стрелкам» показаны главные причинные связи. (С изменениями нз Keith, 1983.)
| Колебания относительных величин биомассы основных компонентов «10-летнего природного цикла» в провинции Альберта, Канада. Стрелкам» показаны главные причинные связи. (С изменениями нз Keith, 1983.) | 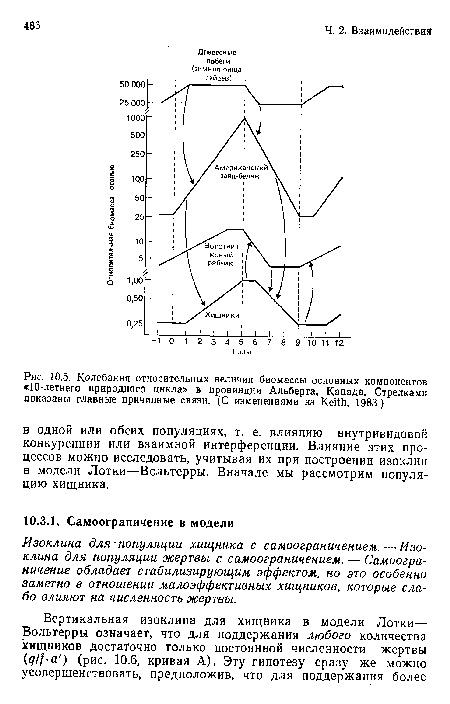 |
Кривыми от А до Г показаны изоклины возрастающей сложности для хищника. А — изоклина по модели Лотки—Вольтерры. Б — для большего числа хищников требуется больше жертв. В — взаимная интерференция между хищниками ведет к прогрессирующему снижению скорости потребления пищи. Г — хищники ограничены не количеством пищи, а другим фактором.
| Кривыми от А до Г показаны изоклины возрастающей сложности для хищника. А — изоклина по модели Лотки—Вольтерры. Б — для большего числа хищников требуется больше жертв. В — взаимная интерференция между хищниками ведет к прогрессирующему снижению скорости потребления пищи. Г — хищники ограничены не количеством пищи, а другим фактором. |  |
Изоклина популяции жертвы с самоограничением, объединенная с изоклинами популяции хищника при возрастающем уровне самоограничения
| Изоклина популяции жертвы с самоограничением, объединенная с изоклинами популяции хищника при возрастающем уровне самоограничения | 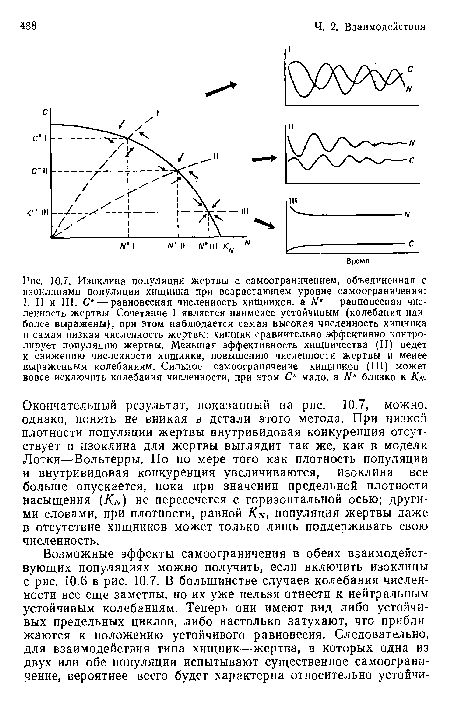 |
Если функциональный ответ типа 3 возникает вследствие переключения хищника на другую жертву, то численность хищника может ие зависеть от плотности жертвы любого типа (основной рисунок) и изоклина хищника может представлять собой горизонтальную линию (не изменяется в зависимости от плотности жертвы). Такая зависимость приводит к устойчивой динамике численности (вверху справа), когда плотность жертвы гораздо ниже предельной плотности насыщения
| Если функциональный ответ типа 3 возникает вследствие переключения хищника на другую жертву, то численность хищника может ие зависеть от плотности жертвы любого типа (основной рисунок) и изоклина хищника может представлять собой горизонтальную линию (не изменяется в зависимости от плотности жертвы). Такая зависимость приводит к устойчивой динамике численности (вверху справа), когда плотность жертвы гораздо ниже предельной плотности насыщения |  |
Изоклина модели хищник—жертва с несколькими равновесными состояниями. А. При низких значениях плотности изоклина жертвы имеет восходящий участок и далее колоколообразиую часть; благодаря этому изоклина хищника может пересекать ее три раза. Пересечения X и 1 представляют собой точки устойчивого равновесия, а пересечение У — это точка неустойчивого положения, из которого значение суммарной численности популяций хищника и жертвы смещается либо к положению X, либо к положению 1. Б. Вероятная траектория, вдоль которой может изменяться суммарная численность популяций, если на нее действуют силы, показанные на рис. А. В. Те же значения численности в зависимости от времени показывают, что пересечение с характеристиками, которые не меняются, может привести к явной «вспышке» численности
| Изоклина модели хищник—жертва с несколькими равновесными состояниями. А. При низких значениях плотности изоклина жертвы имеет восходящий участок и далее колоколообразиую часть; благодаря этому изоклина хищника может пересекать ее три раза. Пересечения X и 1 представляют собой точки устойчивого равновесия, а пересечение У — это точка неустойчивого положения, из которого значение суммарной численности популяций хищника и жертвы смещается либо к положению X, либо к положению 1. Б. Вероятная траектория, вдоль которой может изменяться суммарная численность популяций, если на нее действуют силы, показанные на рис. А. В. Те же значения численности в зависимости от времени показывают, что пересечение с характеристиками, которые не меняются, может привести к явной «вспышке» численности | 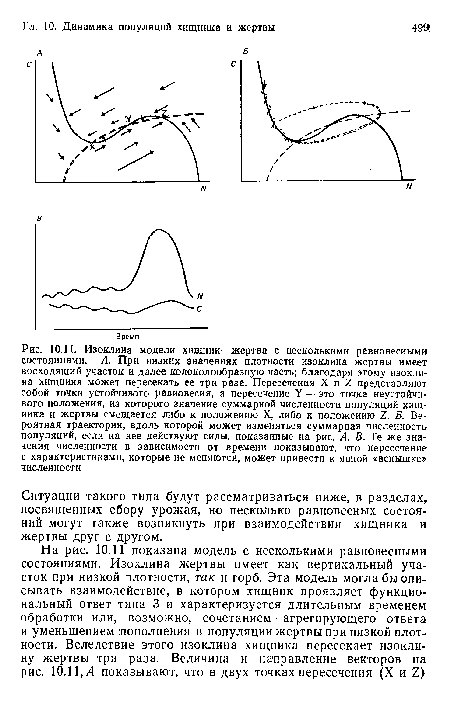 |
Изменения численности цапель (Ardea cinerea) в Англии и Уэльсе (определялась по числу занятых гнезд) хорошо согласуются с изменениями условий среды (особенно суровыми зимами). (Из Stafford, 1971.)
| Изменения численности цапель (Ardea cinerea) в Англии и Уэльсе (определялась по числу занятых гнезд) хорошо согласуются с изменениями условий среды (особенно суровыми зимами). (Из Stafford, 1971.) |  |
Снижение численности популяций антарктических усатых китов под влиянием промысла. (По ОиИапё, 1971.)
| Снижение численности популяций антарктических усатых китов под влиянием промысла. (По ОиИапё, 1971.) | 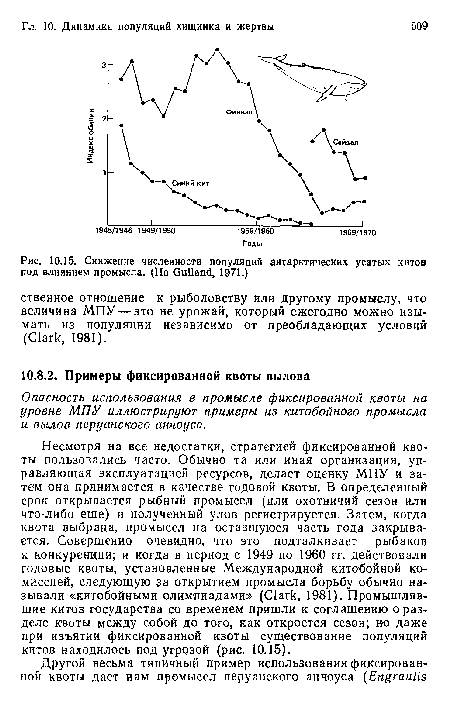 |
Снижение запаса североморской сельди (оценен в миллионах тонн). (Из lies, 1981.)
| Снижение запаса североморской сельди (оценен в миллионах тонн). (Из lies, 1981.) | 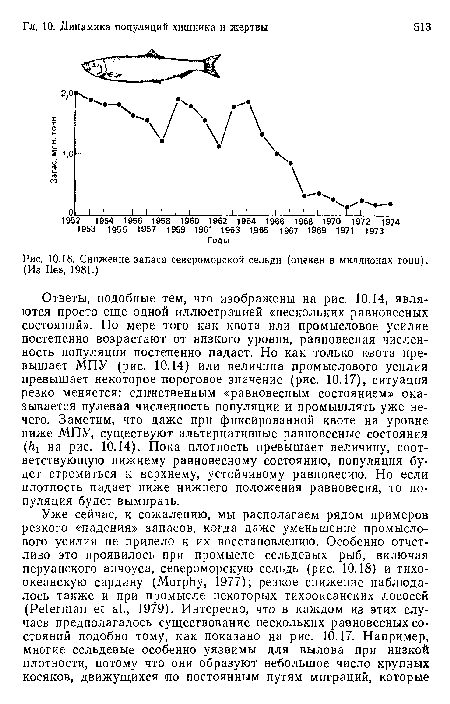 |
Подход к промыслу и управлению рыбными ресурсами на основе объединенного фонда, изображенный в виде блок-схемы. В ее состав входят четыре основные «субмодели»
| Подход к промыслу и управлению рыбными ресурсами на основе объединенного фонда, изображенный в виде блок-схемы. В ее состав входят четыре основные «субмодели» | 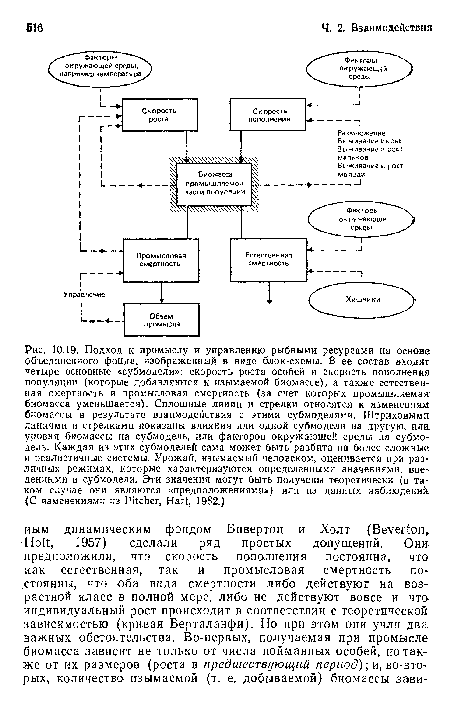 |
Прогнозы Гарро и Джонса (Garrod, Jones, 1974) для стада сайки при трех режимах интенсивности промысла и трех размерах ячеи сети. (По Pitcher, Hart, 1982.)
| Прогнозы Гарро и Джонса (Garrod, Jones, 1974) для стада сайки при трех режимах интенсивности промысла и трех размерах ячеи сети. (По Pitcher, Hart, 1982.) | 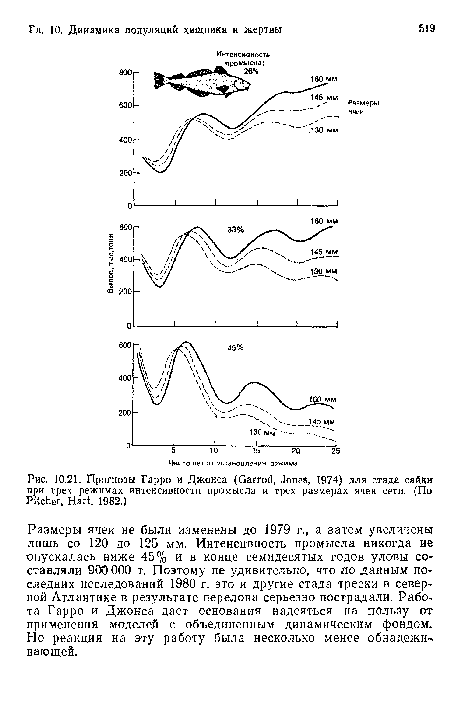 |
Классификация организмов по ширине тела из пищевых сетей наземных редуцентов. Полностью хищными являются следующие группы
| Классификация организмов по ширине тела из пищевых сетей наземных редуцентов. Полностью хищными являются следующие группы | 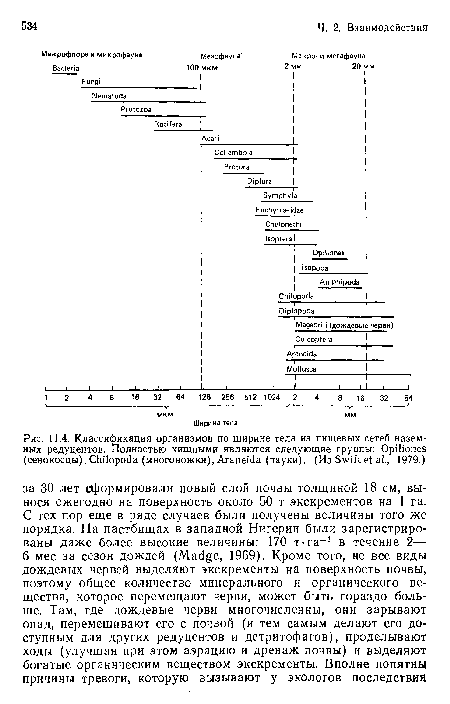 |
Представители наиболее важных групп детритофагов в листовом опаде и почве (масштаб не соблюден)
| Представители наиболее важных групп детритофагов в листовом опаде и почве (масштаб не соблюден) |  |
Примеры различных категорий беспозвоночных консументов в пресных водах.
| Примеры различных категорий беспозвоночных консументов в пресных водах. | 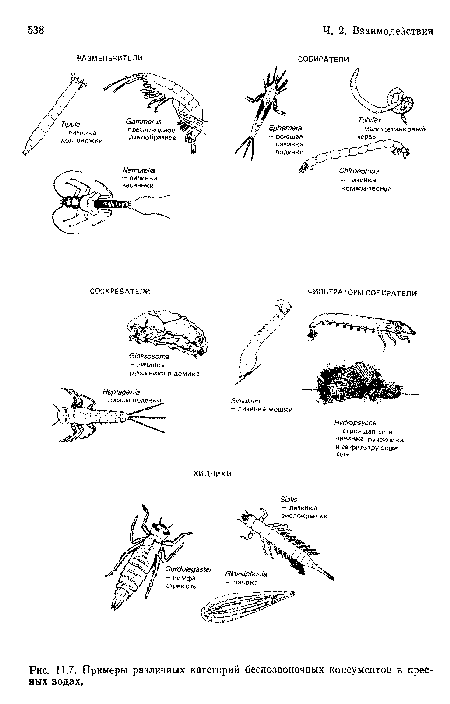 |
Общая схема потока энергии в ручье. Часть крупнозернистой органической взвеси (КОВ) вследствие выщелачивания быстро теряется в виде растворенного органического вещества (РОВ). Оставшаяся часть превращается в тонкую органическую взвесь (ТОВ) благодаря трем процессам
| Общая схема потока энергии в ручье. Часть крупнозернистой органической взвеси (КОВ) вследствие выщелачивания быстро теряется в виде растворенного органического вещества (РОВ). Оставшаяся часть превращается в тонкую органическую взвесь (ТОВ) благодаря трем процессам | 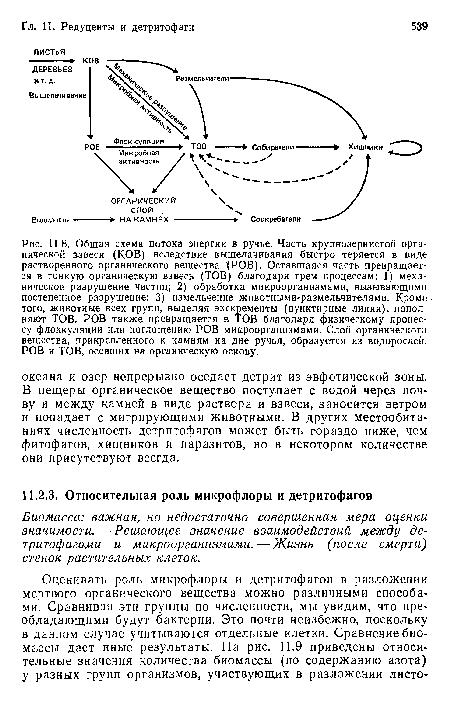 |
Относительное значение микрофлоры в разложении лесного опада по сравнению с членистоногими, дождевыми червями и нематодами, оцененное по содержанию в них азота. Микробная активность гораздо выше, чем активность детритофагов, ио обилие
| Относительное значение микрофлоры в разложении лесного опада по сравнению с членистоногими, дождевыми червями и нематодами, оцененное по содержанию в них азота. Микробная активность гораздо выше, чем активность детритофагов, ио обилие | 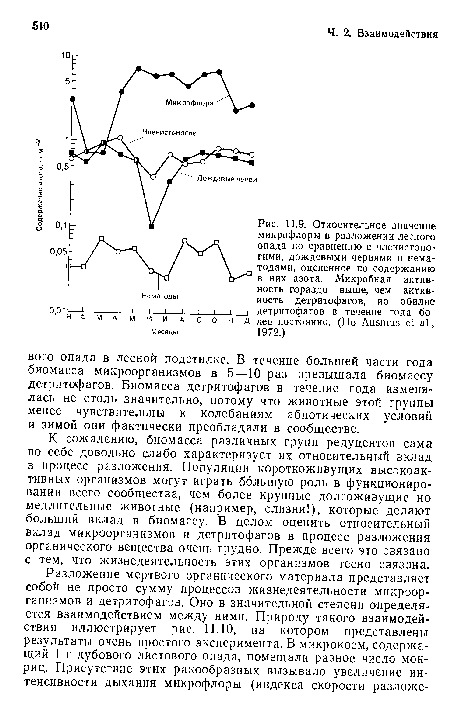 |
Африканский жук-иавозиик, скатывающий шарик из помета (фотография
| Африканский жук-иавозиик, скатывающий шарик из помета (фотография |  |
Скорость разложения трупов мелких млекопитающих в сельской местности графства Оксфорд в разные периоды
| Скорость разложения трупов мелких млекопитающих в сельской местности графства Оксфорд в разные периоды | 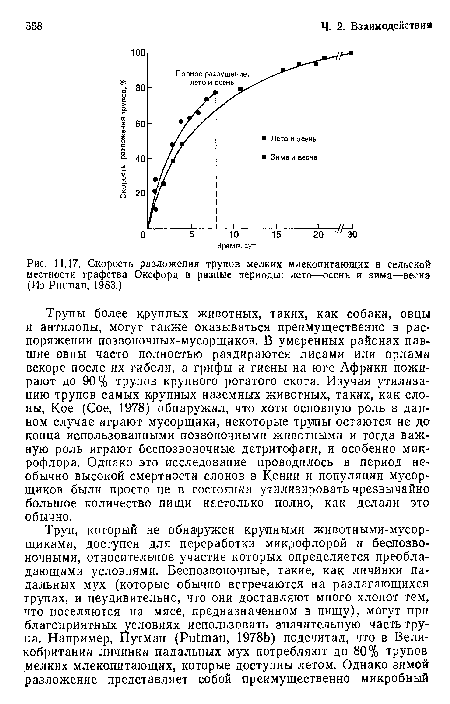 |
Захоронение мыши парой жуков-мертвоедов Necrophorus. (Из Milne, Milne, 1976.)
| Захоронение мыши парой жуков-мертвоедов Necrophorus. (Из Milne, Milne, 1976.) | 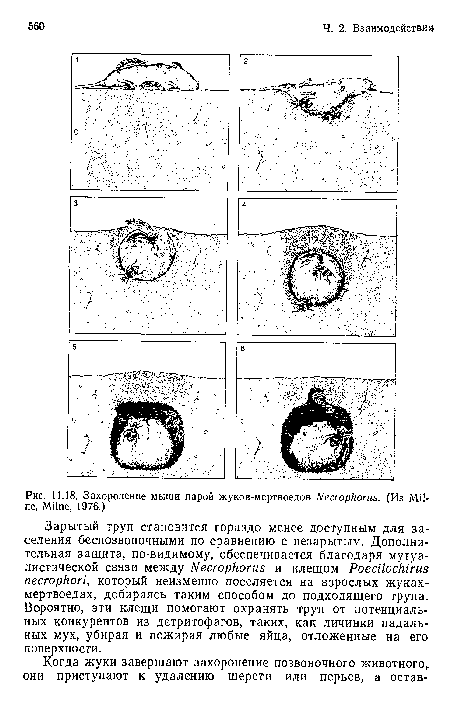 |
Микропаразиты животных. Слева
| Микропаразиты животных. Слева |  |
Жизненный цикл семи наиболее важных паразитов — филярий человека. Филярии — это долгоживущие черви из нематод, которые переносятся кровососущими хозяевами. (По Whitfield, 1982.)
| Жизненный цикл семи наиболее важных паразитов — филярий человека. Филярии — это долгоживущие черви из нематод, которые переносятся кровососущими хозяевами. (По Whitfield, 1982.) | 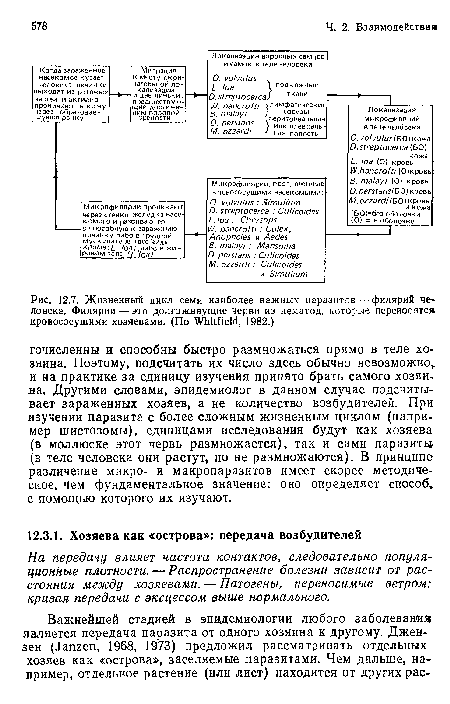 |
Связь между частотой встречаемости и средней интенсивностью зараженности для агрегированного, случайного и регулярного размещений паразитов у хозяев. (По Anderson, 1982.)
| Связь между частотой встречаемости и средней интенсивностью зараженности для агрегированного, случайного и регулярного размещений паразитов у хозяев. (По Anderson, 1982.) | 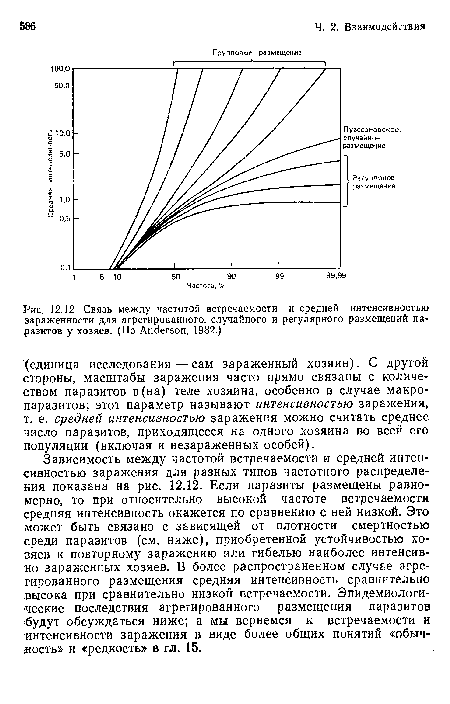 |
Высшее растение представляет собой совокупность специализированных ниш, занятых болезнетворными грибами, хищными и паразитическими насекомыми. Сходную, хотя и менее разветвленную, диаграмму можно было бы составить для нематод и патогенных бактерий. Степень специализации по нишам на самом деле даже выше, чем показано на диаграмме
| Высшее растение представляет собой совокупность специализированных ниш, занятых болезнетворными грибами, хищными и паразитическими насекомыми. Сходную, хотя и менее разветвленную, диаграмму можно было бы составить для нематод и патогенных бактерий. Степень специализации по нишам на самом деле даже выше, чем показано на диаграмме | 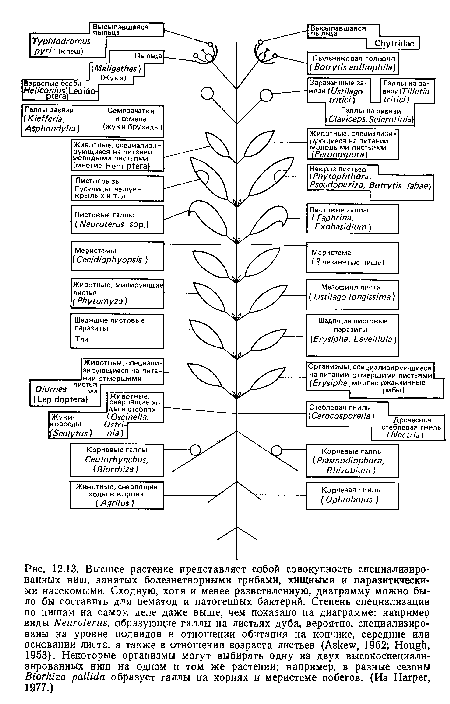 |
Позвоночное животное как хозяин является реагирующей средой, в которой иммуииый ответ превращает его из пригодного для паразита место-обитания в непригодное. Этот тип реакции позволяет отличить живые организмы, которые служат местообитаниями, от тех категорий среды обитания, которые обычно рассматриваются экологами.
| Позвоночное животное как хозяин является реагирующей средой, в которой иммуииый ответ превращает его из пригодного для паразита место-обитания в непригодное. Этот тип реакции позволяет отличить живые организмы, которые служат местообитаниями, от тех категорий среды обитания, которые обычно рассматриваются экологами. | 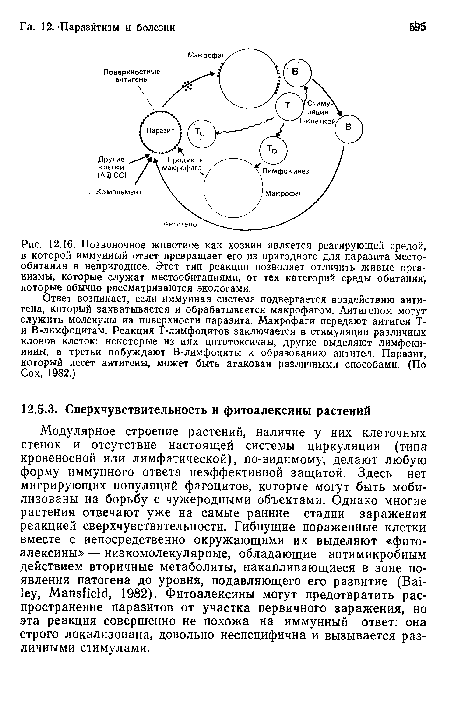 |
Мухи-агромизиды, паразитирующие и образующие галлы иа дубе (см. фото 2).
| Мухи-агромизиды, паразитирующие и образующие галлы иа дубе (см. фото 2). |  |
Зарегистрированные случаи кори в Нью-Йорке с 1944 по 1964 гг. (По Yorke, London, 1973.)
| Зарегистрированные случаи кори в Нью-Йорке с 1944 по 1964 гг. (По Yorke, London, 1973.) |  |
Влияние патогенности паразита на степень подавления им популяции хозяина. А. Модели временной инфекции, в которых зараженные хозяева после заражения не выздоравливают. В заштрихованной области паразиты вымирают и популяция хозяев достигает предельной плотности насыщения К а — скорость размножения хозяина; ¡5 — зараженность и а — вызванная паразитами смертность. На этом рисунке приведены две скорости размножения хозяина
| Влияние патогенности паразита на степень подавления им популяции хозяина. А. Модели временной инфекции, в которых зараженные хозяева после заражения не выздоравливают. В заштрихованной области паразиты вымирают и популяция хозяев достигает предельной плотности насыщения К а — скорость размножения хозяина; ¡5 — зараженность и а — вызванная паразитами смертность. На этом рисунке приведены две скорости размножения хозяина | 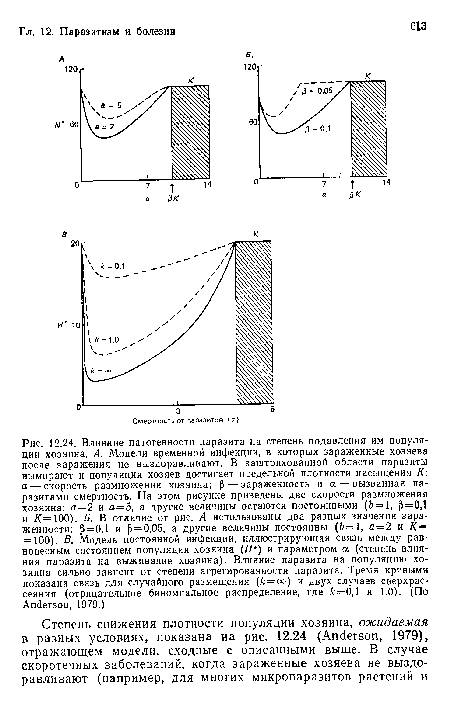 |
Схема, иллюстрирующая варианты взаимодействия генов устойчивости (#) хозяина с пятью соответствующими генами вирулентности паразита в системе циклического сбалансированного полиморфизма. (По Person, 1966.)
| Схема, иллюстрирующая варианты взаимодействия генов устойчивости (#) хозяина с пятью соответствующими генами вирулентности паразита в системе циклического сбалансированного полиморфизма. (По Person, 1966.) |  |
Колебания численности трипаносом в крови больного сонной болезнью. При каждом подъеме численности популяция трипаносом отличается по антигенам от популяции на соседних подъемах. (По Ross, Thomson, 1910.)
| Колебания численности трипаносом в крови больного сонной болезнью. При каждом подъеме численности популяция трипаносом отличается по антигенам от популяции на соседних подъемах. (По Ross, Thomson, 1910.) | 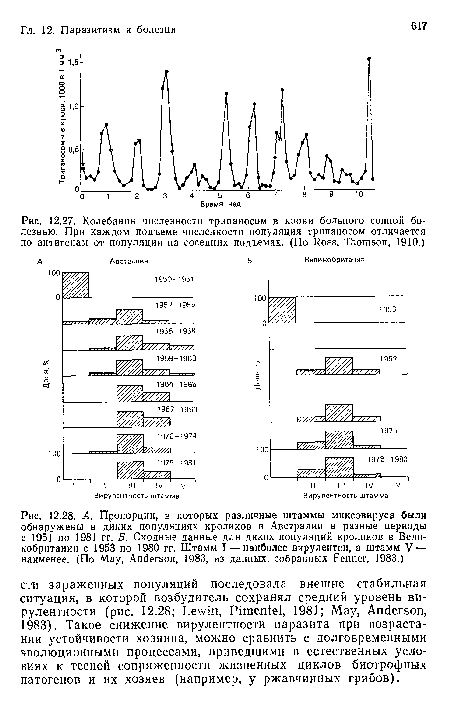 |
А. Пропорции, в которых различные штаммы миксовируса были обнаружены в диких популяциях кроликов в Австралии в разные периоды с 1951 по 1981 гг. Б. Сходные данные для диких популяций кроликов в Великобритании с 1953 по 1980 гг. Штамм I — наиболее вирулентен, а штамм V — наименее. (По May, Anderson, 1983, из данных, собранных Fenner, 1983.)
| А. Пропорции, в которых различные штаммы миксовируса были обнаружены в диких популяциях кроликов в Австралии в разные периоды с 1951 по 1981 гг. Б. Сходные данные для диких популяций кроликов в Великобритании с 1953 по 1980 гг. Штамм I — наиболее вирулентен, а штамм V — наименее. (По May, Anderson, 1983, из данных, собранных Fenner, 1983.) | |
Типы морфологической интеграции между симбионтами и хозяевами А. Симбионты (черные кружки) и организм-хозяин (квадраты)
| Типы морфологической интеграции между симбионтами и хозяевами А. Симбионты (черные кружки) и организм-хозяин (квадраты) | 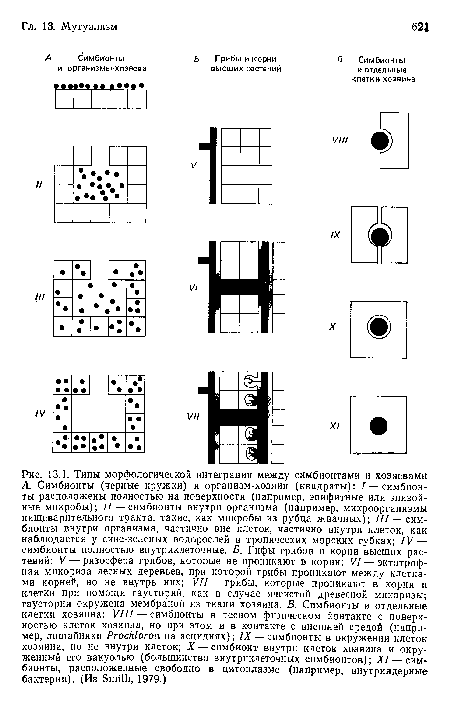 |
Микробное переваривание в рубце овцы. Данные получены при непрерывном питании люцерной. Для использования веществ и веществ, ежедневно образующихся в рубце, получена следующая эмпирическая формула
| Микробное переваривание в рубце овцы. Данные получены при непрерывном питании люцерной. Для использования веществ и веществ, ежедневно образующихся в рубце, получена следующая эмпирическая формула | 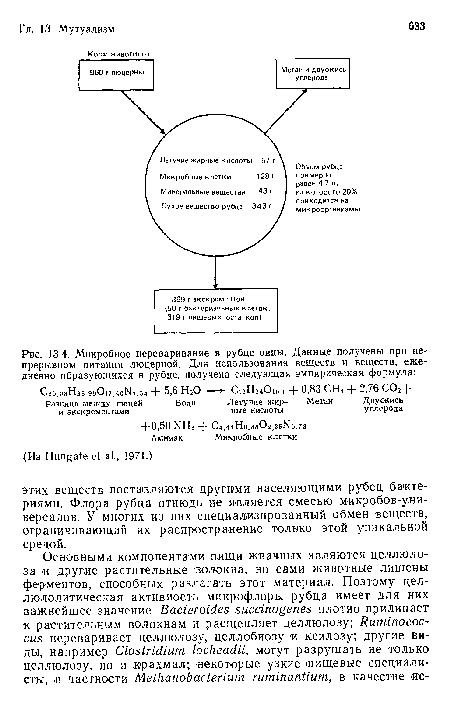 |
Приспособление, использованное для изучения взаимодействия между мутуалистами, в котором партнеры находятся в отдельных камерах, связанных через циркулирующую жидкость. Перемешивание популяций предотвращается с помощью пучков волокон и диализных мембран. (Из Taylor, 1975.)
| Приспособление, использованное для изучения взаимодействия между мутуалистами, в котором партнеры находятся в отдельных камерах, связанных через циркулирующую жидкость. Перемешивание популяций предотвращается с помощью пучков волокон и диализных мембран. (Из Taylor, 1975.) | 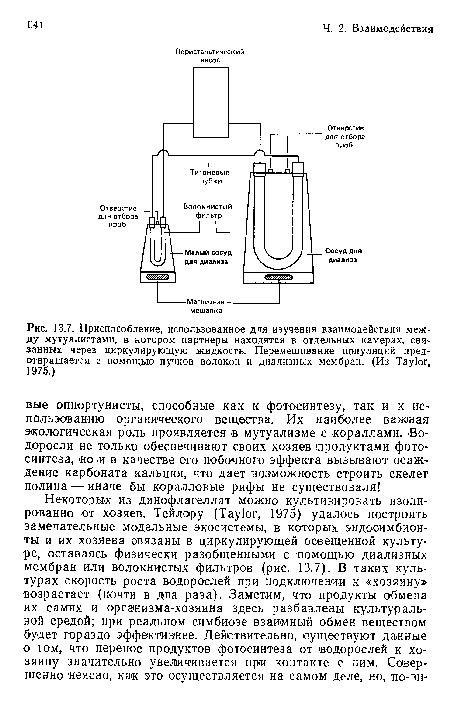 |
Строение некоторых лишайников. А. Простой двухкомпонентный лишайник, например СоИета, в котором клетки водоросли рассредоточены в теле гриба. Б. Более упорядоченная структура, в которой водоросли сосредоточены в центральном слое. В. Срез через специализированное образование (це-фалоподиум), которое служит для расселения и содержит клетки сине-зеленых водорослей.
| Строение некоторых лишайников. А. Простой двухкомпонентный лишайник, например СоИета, в котором клетки водоросли рассредоточены в теле гриба. Б. Более упорядоченная структура, в которой водоросли сосредоточены в центральном слое. В. Срез через специализированное образование (це-фалоподиум), которое служит для расселения и содержит клетки сине-зеленых водорослей. | 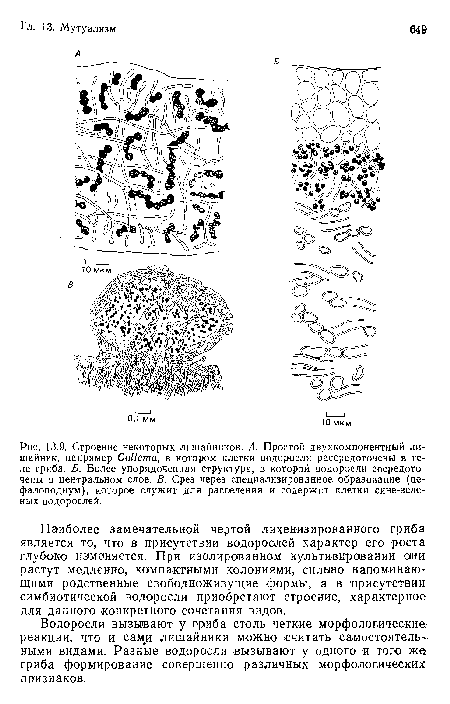 |
Развитие корневого клубенька в результате заражения корня бобового растения клубеньковыми бактериями. (Из Зргеп!;, 1979.)
| Развитие корневого клубенька в результате заражения корня бобового растения клубеньковыми бактериями. (Из Зргеп!;, 1979.) | 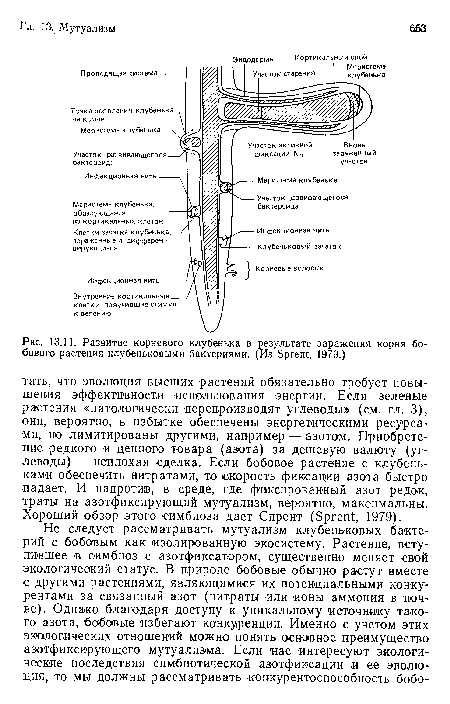 |