Регуляция образования белков-ферментов может осуществляться пе только благодаря непосредственному влиянию репрессоров на молекулы ДНК. Интересны исследования, показывающие, что под влиянием разлпчпы.х воздействий меняется активность РНК-полимера-зы — фермента, катализирующего образование и-РНК на ДНК-мат-рице. Изменение активности этого фермента может оказать влияние на скорость процесса транскрипции. Следующим регуляторным механизмом может быть степень выхода уже образованных и-РНК из ядра. В случае, если под влиянием определенных воздействии поры в оболочке ядра закрываются, ото может тормозить или даже прекращать выход п-РНК из ядра в цитоплазму, что, естественно, приостанавливает синтез белков-ферментов. Для того чтобы служить матрицей, и-РНК должна освободиться от окружающего ее белка. По-видимому, и этот процесс может меняться в зависимости от условий (регуляции па уровне трансляции). Наконец, количество образовавшихся идентичных белков может регулироваться путем изменения времени жизни и-РНК. Рассмотренные иримеры показывают различные возможности регуляции новообразования белков-фермеп-тов. Благодаря этой регуляции организм может приспосабливаться к различным условиям среды. В основе изменении обмена, происходящих под влиянием внесших воздействий, лежит прежде всего изменение направленности и скорости ферментативных реакций. Регуляция образования и активности белков-ферментов лежит также в основе процессов дифференциации.[ ...]
Регуляция синтеза белков-фермептов может происходить разными путями. Работами Жакоба и Mono установлено, что в клетке па-ряду со структурными генами, которые определяют структуру белка, имеются гены-регуляторы, которые ответственны за активацию или дезактивацию работы структурных генов. Предложенная Жакобом и Mono схема регуляции синтеза белка представлена на рисунке 18. Согласно этой схеме гены-регуляторы пространственно отделены от структурных генов и вырабатывают и-РИК, на которой, как на матрице, синтезируются определенные белки (репрессоры). Эти белки-реп-рессоры могут присоединяться к участкам молекулы ДНК, носящим название операторов. В результате на определенном отрезке, включающем, как правило, несколько генов, молекула ДНК перестает раскручиваться, а следовательно, на ней невозможно образование соответствующих и-РНК. Участок ДНК, работа которого регулируется одним оператором, носит название оперона.[ ...]
Внутренняя регуляция. Процессы, протекающие в клетках, подвержены регуляции. На молекулярном уровне регуляторные механизмы существуют в виде обратных химических реакций, основу которых составляют реакции с участием ферментов, обеспечивающие замкнутость процессов регуляции по схеме синтез — распад — ресинтез. Синтез белков, включая ферменты, регулируется с помощью механизмов репрессии, индукции и позитивного контроля. Напротив, регуляция активности самих ферментов происходит по принципу обратной связи, заключающейся в ингибировании конечным продуктом. Известно также регулирование путем химической модификации ферментов. В регуляции активности клеток принимают участие гормоны, обеспечивающие химическую регуляцию.[ ...]
| Схема регуляция синтеза белка. | 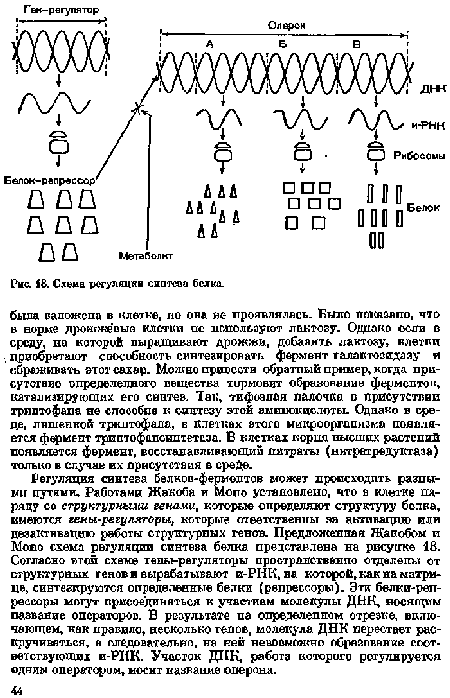 |
Ядро — это центр регуляции жизнедеятельности клетки. Ядро отделено от цитоплазмы ядерной мембраной, в которой имеются поры. Внутри оно заполнено кариоплазмой, в которой находятся молекулы ДНК, обеспечивающие передачу наследственной информации. Здесь происходит синтез ДНК, РНК, рибосом. Часто в ядре можно увидеть одно или несколько темных округлых образований — это ядрышки. Здесь образуются и скапливаются рибосомы. В ядре молекулы ДНК не видны, так как находятся в виде тонких нитей хроматина. Перед делением ДНК спирализуются, утолщаются, образуют комплексы с белком и превращаются в хорошо заметные структуры —хромосомы (рис. 49). Обычно хромосомы в клетке парные, одинаковые по форме, величине и наследственной информации. Парные хромосомы называются гомологичными. Двойной парный набор хромосом называется диплоидным. В некоторых клетках и организмах содержится одинарный, непарный набор, который называется гаплоидным.[ ...]
Гормональная регуляция физиологических процессов у растений па уровне синтеза PIIK и белка.[ ...]
Приспособительные изменения белков могут быть как количественными, так и качественными, и касаются они прежде всего ферментных белков — двигателей жизни. Известны четыре типа таких изменений: увеличение или уменьшение количества и концентрации белковых макромолекул в результате усиления или угнетения их синтеза; присоединение к существующим макромолекулам различных веществ, модифицирующих высшие белковые структуры, а значит, и функциональные свойства белков (в случае белков-ферментов это всякие активаторы и ингибиторы); изменения регуляции функций того или иного белка; построение нового типа макромолекул. Обратимся к конкретным примерам.[ ...]
Считают также, что у эукариотов в генетической регуляции имеет значение структура хроматина, которая блокирует доступ специфических активирующих белков (активаторов) к промоторам. В клетках эукариотов установлен специфический комплекс, который обеспечивает АТФ-зависимое разрушение нуклеосом, позволяя при этом активаторам связываться с нуклеосомным стержнем, что ведет к транскрипции. С другой стороны, установлено, что синтез некоторых ферментов интенсифицируется под влиянием стероидных гормонов. Контроль может осуществляться и после трансляции (контроль протеолиза).[ ...]
| Схема, иллюстрирующая теорию регуляции синтеза белка Жакоба |  |
Много данных и о связи действия цитокининов с синтезом белка. Обработка кинетином стареющих листьев не только задерживает распад белка, но и увеличивает его образование. Показано, что под влиянием цитокиинпов возрастает количество всех видов РНК. Воз можно влияние цитокининов на уровне транскрипции. В исследовани ях 0. Н. Кулаевой с сотрудниками показана прямая стимуляция но], влиянием цитокинипа активности выделенного хроматина. Поскольку новообразование белков-ферментов закодировано в генбме клетки, то это позволяет предположить осуществление гормональной регуляции на уровне гена (транскрипция) или на уровне рибосом (трансляция). Возможно, что фитогормоны, подобно гормонам животных организмов, регулируют дифференциальную активность генома и тем самым влияют на новообразование соответствующих белков-ферментов.[ ...]
Метаболизм подвержен непрерывной генетической регуляции, которая осуществляется в клетках путем регуляции синтеза ферментов. В клетках синтезируется столько малых молекул, сколько их необходимо для синтеза нуклеиновых кислот, белков, липидов и углеводов. Это позволяет оценивать клетки в качестве чрезвычайно экономичных саморегулируемых химических систем.[ ...]
Об этом же косвенно свидетельствуют данные о содержании белка в кишечнике и мышцах молоди леща, полученные при помощи разных методов. Известно, что метод Лоури основан на определении в молекулах белка количества ароматических аминокислот (преимущественно тирозина). Также известно, что тирозин у позвоночных животных является предшественником синтеза гормонов щитовидной железы (тироксина), гормонов мозгового слоя надпочечников (адреналина) и меланинов. В свою очередь синтез пищеварительных ферментов находится под гормональным контролем. Особо следует отметить, что тироксин выполняет ряд функций, в том числе увеличивает энергетический обмен, рост и дифференцировку тканей. У костистых рыб, кроме того, тироксин стимулирует метаморфоз. Недавно была высказана гипотеза, согласно которой основная роль тиреоидных гормонов у рыб заключается в регуляции преадаптаций животных за счет изменений в кишечнике и других структурах, связанных с начальными этапами ассимиляции пищи [22]. Отсюда ясна исключительно важная роль тирозина на ранних этапах онтогенеза костистых рыб.[ ...]
Большое число работ было посвящено изучению влияния ростовых веществ на синтез РНК и белка в семенах, правда, полученные результаты оказались весьма различными и нередко противоречивыми. По некоторым данным, АБК ингибирует синтез РНК и ферментов, а ГА3 или кинетин снимают этот эффект в определенных системах. В зародышах груши АБК ингибирует активность РНК-полимеразы, но этот эффект снимается кинетином и ГА3. Образование мРНК для «-амилазы в клетках алейронового слоя ячменя происходит в ответ на действие ГА3 (с. 149), но этот эффект ингибируется АБК. В других работах указывается, что ГА3 увеличивает доступность ДНК-матрицы для транскрипции в семенах лещины. Значение этих наблюдений в связи с гормональной регуляцией покоя и прорастания недостаточно ясно.[ ...]
Кроме неорганических веществ в клетке присутствуют и органические вещества: белки, липиды (жиры), углеводы (сахара), нуклеиновые кислоты (ДНК, РНК). Они составляют основную массу клетки. Наиболее важными органическими веществами являются нуклеиновые кислоты и белки. Нуклеиновые кислоты (ДНК и РНК) участвуют в передаче наследственной информации, синтезе белков, регуляции всех процессов жизнедеятельности клетки.[ ...]
При адаптации организма к условиям среды немалое значение принадлежит гормональной регуляции обмена веществ и физиологических функций. Но этим дело не ограничивается. К приспособительным реакциям организма имеют отношение гормоны щитовидной железы, инсулин — гормон островков поджелудочной железы, соматотропный гормон гипофиза, стероидные андрогены, выделяемые корой надпочечников и мужскими половыми железами, и мн. др. Но все они являются не непосредственными деятелями приспособления организма, а регуляторами его молекулярных основ. Они не производят, а способствуют приспособительной реакции обмена веществ, создавая для нее необходимый «фон». Действие гормонов может быть непосредственным, но чаще имеет ряд посредников. Фосфорилированный гистон, отщепляясь от ДНК, делает транскрипцию возможной.[ ...]
Явление индукции и репрессии ферментов привело Жакоба и Моно к созданию общей теории регуляции активности генов у бактерий, которую мы сейчас кратко рассмотрим. Геи, определяющий структуру какого-либо специфического ферментного белка, называют структурным геном. Синтез мРНК начинается только в определенном локусе на цепи ДНК, получившем название оператора (рис. 13.6). В некоторых случаях один оператор может регулировать транскрипцию нескольких соседних структурных генов. ДНК, находящийся под контролем одного оператора, называют опероном. Ои может содержать один или несколько структурных генов.[ ...]
Несомненно, что для нормального длительного роста растяжением необходим непрерывный синтез белка, даже если краткосрочные (с лаг-периодом порядка 10 мин или меньше) ростовые ответы на гормон наблюдаются и без синтеза новых РНК и белков. В случае регуляции роста растяжением ауксином очевидно, что при быстром ростовом ответе происходит выделение И+ в оболочку, а снижение pH или непосредственно ослабляет какие-то межмолекулярные связи, или благоприятствует активности определенных ферментов, разрыхляющих оболочку.[ ...]
В опытах по выяснению взаимодействия фитогормопов и ингибиторов нуклеинового и белкового синтеза были вскрыты некоторые пути их действия на сексуализацию растений через изменение активности генетического аппарата. Однако имеются и другие подходы к выяснению механизмов регуляции проявления пола па генном уровне. Можно получить такие данные путем выявления различий в белковых спектрах мужских и женских растений. В литературе имеются сведения о том, что листья мужских п женских растений конопли Mercurialis апппа имеют разный состав белков-антигенов [Durand В., Durand R., 1979].[ ...]
До сих пор мы говорили о развитии как о процессе «избирательной экспрессии генов», при которой происходит регуляция активности специфических групп генов, в свою очередь регулирующих синтез ферментов и структурных белков, характерных для специализированных клеток. Информация, закодированная в ДНК, определяет последовательность аминокислот в полипсп-тидиых цепях белков, т. е. их первичную структуру. Образование же структур более высоких порядков зависит от первичной структуры полипептидиой цепи и не нуждается в регуляции со-стороны генома. Иными словами, генетический контроль первичной структуры определяет вторичную, третичную и четвертичную структуры белка.[ ...]
Биологическое значение повторов в «эгоистической» ДНК также не выяснено. Однако предполагают, что они вовлечены в регуляцию экспрессии и рекомбинацию генов, а также в «защиту» некоторых структурных генов, в частности генов, детерминирующих синтез гистонов, рРНК или рибосомных белков.[ ...]
Верна ли гипотеза Бриттена и Дэвидсона, покажет будущее, но работа, проведенная на растениях табака, соответствует гипотезе посттраискрипционной регуляции экспрессии генов. Так, только 75% последовательностей мРНК листьев содержится в полисомах стебля, но в ядерной РНК стебля имеются все мРНК, характерные для листьев табака. Таким образом, структурные гены листьев, неиспользуемые для синтеза белков стебля, транскрибируются в ядре стебля. Это говорит о том, что «посттранскрипционный процессинг, или механизм отбора, играет важную роль в регуляции экспрессии генов у растений» (Голдберг).[ ...]
Исследования молекулярных механизмов в этнологии генных болезней позволило установить существование таких болезней, возникновение которых связано с нарушениями регуляции действия генов.[ ...]
В кратком выражении эта цепь может быть представлена таким образом: внешние воздействия — изменения в метаболизме — влияние на активность генетического аппарата — синтез: специфических РНК и белков — цветение. И если раньше как основные движущие силы, подводящие к переходу растений к цветению, утверждались гормональные вещества, позднее трофические и гормональные факторы, а затем гормональная и генетическая регуляция, то теперь мы подходим к утверждению о томг что такими движущими силами зацветания растений являются все три взаимодействующие системы — гормональная, трофическая и генетическая.[ ...]
Жизнедеятельность организма регулируется прежде всего на субклеточном и молекулярном уровнях. Это химическая авторегуляция реакций обмена веществ. Она решает местные задачи и является основой всех видов регуляции. Осуществляется она путем изменения концентраций метаболитов, повышения или снижения активности н количественного содержания ферментов, т. е. усиления или угнетения их синтеза, структурных изменений их и других функциональных белков. Но регуляция происходит и на более высоких уровнях: клетки в целом, ткани, органа, функциональной системы, организма. Чем на более высокий уровень передаются управляющие выходные сигналы, тем более обобщенный характер они носят. У человека и животных высшим центром, управляющим вегетативными функциями (кровообращением, дыханием, движением, выделением гормонов и т. п.), является гипоталамус, расположенный в нижней части промежуточного мозга, имеющий связи с системой желез внутренней секреции, другими частями мозга и центром сознания — его корой. Поступающие сигналы могут осознаваться или не осознаваться. Управляющие ответы на неосознанные сигналы среды могут осуществляться гипоталамусом и без участия высшего отдела головного мозга — его коры.[ ...]
Ацетилхолин и биогенные амины могут регулировать активность ферментов или включаться в метаболизм как субстраты.[ ...]
В случае ДНК-содержащих вирусов человека и животных их свойство вызывать опухоли зависит от отношения вирусной ДНК к хромосомам клетки. Вирусная ДНК может оставаться подобно плазмидам в клетке в автономном состоянии, реплицируясь вместе с клеточными хромосомами. При этом регуляция деления клеток не нарушается. Однако вирусная ДНК может включиться в одну или несколько хромосом клетки-хозяина. При таком исходе деление клеток становится нерегулируемым. Другими словами, инфицированные ДНК-содержащим вирусом клетки превращаются в раковые. Примером онкогенных ДНК-содержащих вирусов является вирус вУ40, выделенный много лет назад из клеток обезьян. Онкогенное действие этих вирусов зависит от того, что отдельные вирусные гены действуют как онкогены, активизируя клеточную ДНК и побуждая клетки к вступлению в в-фазу с последующим неконтролируемым делением. РНК-содержащие вирусы из-за включения их РНК в одну или несколько хромосом клетки-хозяина также обладают онкогенным действием. В геноме этих вирусов также есть онкогены, однако они существенно отличаются от онкогенов ДНК-содержащих вирусов тем, что в геноме кле-ток-хозяев присутствуют их гомологи в виде протоонкогенов. Когда РНК-содержащие вирусы инфицируют клетки, они «захватывают» в свой геном протоонкогены, которые представляют собой последовательности ДНК, контролирующие синтез белков (киназ, факторов роста, рецепторов факторов роста и др.), участвующих в регуляции клеточного деления. Однако известно, что существуют и другие способы превращения клеточных протоонкогенов в вирусные онкогены.[ ...]
Клетка состоит из наружной клеточной мембраны, цитоплазмы с органеллами и ядра. Животные и растительные клетки в своем строении имеют особенности. Некоторые из составных элементов клеток, например как хлоропласты и вакуоли, характерны только для растительных клеток, которые имеют кроме мембраны еще и оболочку (рис. 25). Важнейшим элементом клетки для осуществления функций самовоспроизведения и наследственности является ядро — центр регуляции жизнедеятельности клетки. Ядро заполнено кариоплазмой, содержащей молекулы ДНК; в ядре происходит синтез ДНК, РНК, рибосом. Перед делением ДНК образуют комплексы с белком, формируя хромосомы (рис. 26). Число хромосом для каждого вида организма постоянно, это собственно и определяет возможность появления живых организмов одного и того же вида в процессе размножения (рис. 27).[ ...]