Кофермент — небелковый органический компонент фермента, ответственный за его активность (Часто витамин).[ ...]
Поведение кофермента в активном центре дегидрогеназ продолжает оставаться одним из наиболее интересных вопросов как с точки зрения взаимного влияния отдельных элементов структуры НАД на реакционную способность никотинамидной компоненты, так и с точки зрения понимания механизма последующего связывания субстрата и влияния на саму белковую молекулу, в частности на ее вторичную, третичную и четвертичную структуры..[ ...]
Эти соединения во многих отношениях отличались от традиционных комплексных соединений трехвалентного кобальта. В связи с этим проблема их исследования включает определение реального валентного состояния атома кобальта, изучение реакционной способности кобальта, выяснение роли групп, замещающих транс-аксиальные лиганды и их влияния на свойства корриновой системы, а также на другие гранс-аксиальные лиганды и на атом кобальта. Исследование химических и физико-химических свойств кобамидного кофермента является основой для понимания механизма действия В -зави-симых ферментов.[ ...]
| Связи кофермента с белком в отсутствие субстратов в активной форме фермента, pH 8,0 [см. схему 1 (1)]. | ![Связи кофермента с белком в отсутствие субстратов в активной форме фермента, pH 8,0 [см. схему 1 (1)].](/static/pngsmall/827885330.png) |
Осуществлению (полного синтеза кофермента А предшествовало получение следующих основных компонентов его молекулы — £>-( + )-пантотеновой кислоты; 0-( + )-пантетеина- и его 4 -фосфата; аденозин-3 ,5 -дифос-фата.[ ...]
Следовательно, коферменты (и синтетические структурные аналоги коферментов) являются прекрасным инструментом для изучения структуры и функции активного центра фермента.[ ...]
История изучения коферментов рёзвивается удивительно неровно. Она переживала и стремительные взлеты, вызывая энтузиазм исследователей, когда казалось, что именно на этом пути механизм ферментативного катализа будет расшифрован. Были и периоды пессимизма, когда оказывалось, что, несмотря на исчерпывающую ясность строения и химизма превращений коферментов в ходе каталитического акта, истинный молекулярный механизм действия ферментов по-прежнему остается «синей птицей». В наше время эта область исследований развивается относительно спокойно и вклад химии и биохимии коферментов в общую проблему ферментативного катализа становится все более ясно очерченным.[ ...]
Истинная история коферментов начинается спустя примерно 30 лет после открытия Harden и Yong и связана в первую очередь с работами лабораторий Warburg и Euler.[ ...]
Анализ структуры коферментов и данных относительно их взаимодействия со специфическими белками позволяют условно выделить в коферментах два функциональных участка —группировку, ответственную за связывание с белком, и собственно каталитическую часть.[ ...]
Исходное состояние кофермента в белке ААТ изображено на рис. 67 и схеме.[ ...]
| Структура ДБК-кофермента, полученная на основании данных рентгеноструктурного анализа. | 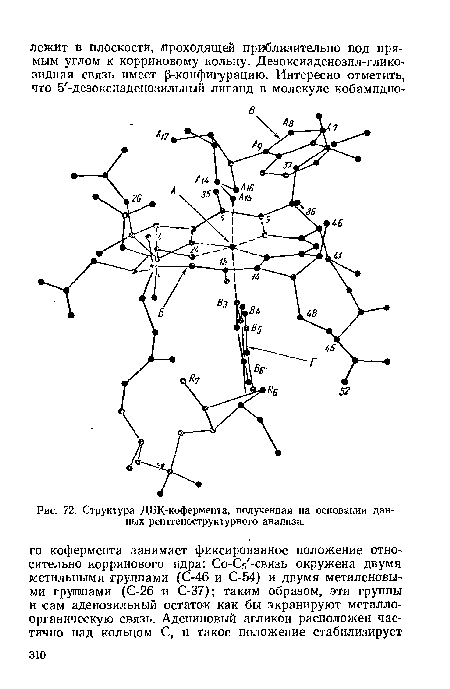 |
Эти соединения представляют собой окрашенные в ярко-желтый цвет вещества, обладающие интенсивной желто-зеленой флюоресценцией, хорошо растворимые в воде, растворимые в пиридине и феноле и не растворимые в большинстве органических растворителей.[ ...]
Установлено, что роль коферментов во многих случаях выполняют различные витамины. В настоящее время прежнее резкое разграничение между ферментами, коферментами и активными труппами постепенно стирается.[ ...]
Острое локальное снижение уровней коферментов является не редким патологическим признаком заболевания, и в этом случае применение готовых коферментов может быть одним из эффективных путей терапии. Это направление использования коферментов вызывает необходимость разработки практически удобных методов их синтеза, исследования фармакологии коферментов, в том числе проблем проникновения их в ткани организма, а также многих других вопросов. А это в свою очередь требует широкого изучения химии и механизма действия коферментов, некоторые результаты которого составляют содержание этой книги.[ ...]
Для НАД остается неизвестной конформация кофермента при его связывании в активном центре дегидрогеназ. Основной вклад в расшифровку этого трудного вопроса, как можно ожидать, будет сделан с помощью таких прямых методов, как рентгеноструктурный анализ и ЯМР. Работы в этом направлении, начатые в 1968—1970 гг. группами Rossman и Kaplan, позволили получить первые данные, свидетельствующие о развернутой структуре НАД в активном центре лактатдегидрогеназы.[ ...]
Было установлено, что функционирование этого кофермента состоит в обратимом окислении — восстановлении изоаллоксазиновой группировки.[ ...]
Макроцикл содержит 9 асимметрических атомов углерода. Все пропионамидные, заместители находятся на той стороне, где нуклеотид, все ацетамидные группы—на противоположной стороне плоскости корри-нового кольца.[ ...]
Активно продолжаются исследования роли структурных элементов кофермента Bi2 в механизме связывания с апоферментом и в каталитическом действии.[ ...]
Однако не только ферменты регулируют процессы метаболизма. Им помогают коферменты — это крупные молекулы, частью которых являются витамины — вещества, необходимые для обмена веществ всех организмов — бактерий, зеленых растений, животных и человека. Отсутствие витаминов ведет к болезням: нарушается обмен веществ.[ ...]
Если это так, то, по-видимому, придется отказаться от единого механизма действия ДБК-кофермента для всех реакций, что маловероятно, принимая во внимание их общую природу. Во всяком случае, вопрос о гетеро- или гемолитическом разрыве Со-С-связи в молекуле кофермента ■под влиянием белка и о том, какова природа активированного кофермента, является кардинальным для понимания общего механизма действия В -зависимых ферментов.[ ...]
В 1931 г. в лаборатории Warburg при изучении окислительного распада углеводов был обнаружен кофермент глю-козо-6-фосфатдегидрогеназы. Этот кофермент в 1933— 1935 гг. был выделен в индивидуальном состоянии и было установлено наличие в его составе амида никотиновой кислоты, аденина, рибозы и фосфорной кислоты. Тогда же в лаборатории Warburg было выяснено, что механизм действия ТПН и ДПН состоит в обратимом окислении — восстановлении пиридинового ядра. В 1936 г. Karrer пришел к заключению, что присоединение водорода в процессе восстановления ТПН происходит по второму углеродному атому пиридинового ядра. Как известно, эта ошибочная точка зрения держалась довольно долго, до 1955 г., когда Loewus, Vennesland и Harris в опытах с дейтерием показали, что в действительности восстановление является стереоспецифичным и идет в положении 4.[ ...]
Дальнейшим этапом исследований свойств ДБК-кофер-мента явилось изучение кривых кругового дихроизма (КД) кофермента и его аналогов. Хотя однозначной интерпретации кривых КД еще не существует, изучение спектров КД различных корриновых соединений показывает, что имеется параллель между кривыми КД и ультрафиолетовыми спектрами. Особенно важным оказалось свойство кривых КД претерпевать инверсию при замещении гранс-аксиаль-ных лигандов X и Y, в то время как на ультрафиолетовые спектры такое замещение оказывает небольшое влияние. Интересными оказались результаты, полученные нами при исследовании кривых КД 5 -дезоксинуклеозидных .аналогов ДБК-кофермента. В этом случае оказалось, что при 300— 600 нм кривые КД-кофермента и аналогов практически идентичны, а в области 230—300 нм в некоторых случаях наблюдается большое различие. Эти результаты безусловно необходимо учитывать в сравнительном изучении кривых КД В -завмсимых ферментов.[ ...]
В случае применения цельноклеточных катализаторов наиболее доступным способом регенерации восстановленных форм коферментов является использование экзогенных соединений, окисляющихся под действием КАБ(Р)+-зависимых дегидрогеназ, имеющихся в клетках (Ма18ис1а, 2002). В связи с этим была предпринята попытка регенерации активности отработанной биомассы путем ее инкубирования в присутствии экзогенных восстановителей (этанола, лактата и сахарозы).[ ...]
Отличительным свойством этих 2 ферментов является быстрая их инактивация в отсутствие субстрата при добавлении ДБК-кофермента к апоферменту [35, 37]. Эта инактивация наблюдается в аэробных условиях и сопровождается структурными изменениями в коферменте с разрывом Со-С-связи и превращением последнего в оксикоба-ламин.[ ...]
Для осуществления реакции необходимо присутствие НАД; в некоторых случаях, как, например, для УДФ-глю-козо-4-эпимераз дрожжей ■ [10] и высших растений [11], этот кофермент достаточно прочно связан с апоферментом. Внутри фермент-субстратного комплекса (X) происходит перенос гидрид-иона с образованием восстановленной формы НАД; последнее доказано на основании изменения спектров флюоресценции фермента в ходе реакции [12]. Одновременно происходит, по-видимому, атака нуклеофильной группы фермента на водород гидроксильной группы или углеродный атом С-4 с образованием связанного с ферментом УДФ-4-кетосахара, как это изображено на формуле XI, или его производного. Следующей стадией процесса является перегруппировка фермент-субстратного комплекса, в итоге которой изменяется относительное положение НДФ-сахара и восстановленного пиридинового нуклеотида.[ ...]
Однако, каталитическая активность фермента не зависит от природы металла комплексообразователя, функция которого, вероятно, состоит в координации при связывании кофермента и апофермента [53].[ ...]
Одним из важных направлений исследования механизма действия дегидрогеназ является изучение природы функциональных групп активного центра, участвующих в связывании кофермента и субстратов и в катализе. Невозможно без точного знания групп, образующих так называемые якорные площадки субстрата и кофермента, а также групп, принимающих участие непосредственно в каталитическом акте, последовательно и достоверно описать многостадийную ферментативную реакцию.[ ...]
Вопрос о связывании и функционировании НАД в активном центре является общим для ряда дегидрогеназ независимо от их субстратной специфичности. Стадия взаимодействия с коферментом является определяющей в том смысле, что предшествует связыванию субстрата, и может быть поэтому исследована независимо в двухкомпонентной системе фермент—кофермент. Рассмотрению данных такого рода в этом обзоре предшествует описание структуры и свойств молекулы НАД, а также характеристика функциональных групп белка — второго участника реакции образования фермент-коферментного комплекса ■.[ ...]
Это имеет место при подкислении растворов кобаламинов.[ ...]
Вся эта система имеет молекулярпую массу 4,0x10®. В ре-зультате этого процесса образуется активный ацетат — ацетилкоэн-гим А (ацетил-КоА), восстановленный НАД (НАД Н+Н+), п выделяется углекислота (первая молекула). Восстановленный НАД но-ступает в цепь переноса электронов, а ацетил-КоА вступает в цикл трпкарбоновых кислот. При яакоплепии АТФ выше определенного уровня превращение пировиноградной кислоты подавляется. Это один из способов регуляции интенсивности протекания аэробной фазы.[ ...]
После опубликования этих данных Со1оиаск писал: «Это сообщение явилось полным сюрпризом для тех из нас, кто воспринимал как догму принцип .прямого переноса водорода с субстрата на кофермент» [98].[ ...]
Интересными представляются данные о роли остатков триптофана в механизме действия дегидрогеназ. Из работ Westheimer, ставших классическими, следовало, что в активном центре дегидрогеназ осуществляется прямой перенос водорода с субстрата на кофермент [49, 93, 94]. Вместе с тем автор не исключал того, что в роли промежуточного переносчика может выступать какая-либо функциональная Группа белка. Ответ на вопрос о таком участии белка в реакции был, казалось, получен в работах Schellenberg [95—97].[ ...]
Приведенные данные окисления и хлорирования показывают, что реакционная способность хромофора в кофермен-тах и сульфонатах кобаламина отличается от реакционной способности циан- и оксикобаламинов. Но заместители при С-10 хромофора в свою очередь оказывают сильное влияние на прочность связи кобальта с лигандами и тем самым на его реакционную способность.[ ...]
Мотонейроны — двигательные нервные клетки спинного мозга МСГ — меланоцитстимулирующий гормон нижнего мозгового придатка (гипофиза), стимулирующий синтез коричневого пигмента меланина в коже и сетчатке глаза НАД — никотинамидадениндинуклеотид — кофермент см. биологического окисления, принимающий водород от окисляемых веществ и передающий его на дыхательную цепь см.[ ...]
В подавляющем большинстве случаев обязательным компонентом фермента является белок с молекулярной массой от десяти тысяч до нескольких миллионов, который либо сам является катализатором индикаторной реакции, либо становится таковым при образовании комплексов с коферментами или ионами металлов При этом скорость ферментативной реакции зависит от природы субстрата и фермента, температуры, состава среды, в которой протекает реакция и др Характер указанных зависимостей различен для разных ферментов и требует специальных исследований при выборе оптимальных условий определений. Кроме того, активность ферментов во многом зависит от присутствия в системе субстрат-фермент эффекторов - веществ, которые ингибируют или ускоряют каталитический процесс. В частности, к ним относятся и суперэкотоксиканты. Круг ферментов достаточно широк, однако практический интерес представляют лишь те, которые имеют высокую специфичность и обеспечивают низкие пределы обнаружения. На практике из большого числа ферментов при определении суперэкотоксикантов внимания заслуживают около 20, причем в иммобилизованном состоянии - 8-10 (87 . Среди них широкое применение находят холинэстеразы, относящиеся к ферментам-гидролазам [84 ].[ ...]
Известно большое число работ, (посвященных функциональным группам дегидрогеназ, однако не все из них носят систематический характер. Известно, что фермент-коферментный комплекс диссоциирует под действием SH-реагентов, а кофермент обладает защитным действием при инактивации п-хлормер-курибензоатом.[ ...]
Недавно были высказаны сомнения по поводу схемы реакций, предложенных Schellenberg (1, 2). О способности остатков триптофана окисляться до иона индоленина, т. е. о существовании восстановленной формы фермента, можно было бы судить по протеканию реакции изотопного обмена в системе аце-тальдегид—этанол (С14) в отсутствие кофермента. Это обнаружено не было в работе Wong, Williams [99]. По-видимому, остатки триптофана не принимают участия в катализе на промежуточных стациях, но связывание кофермента и субстрата приводит к лабилиза-ции триптофана, в результате которой при денатурации и выделении белка происходит включение метки в метиленовую группу триптофана. Эти результаты согласуются с данными Kaplan, не обнаружившего ферментативный обмен между окисленной и восстановленной формами кофермента в отсутствие субстратов [100].[ ...]
К 1-й группе относятся реакции трансметилирования, требующие витамина Bi2 или одного из его аналогов. Общим для этих реакций является образование Со-метилкор-риноидов как промежуточных переносчиков метальной группы. Ко 2-й группе относятся реакции, катализируемые ферментами, простетической группой которых является кобамидный кофермент (б -дезоксиаденозилкобаламин, ДБК-кофермент). Для этих реакций общим является перенос водорода и образование новой углерод-водородной связи.[ ...]
Особенно важно подчеркнуть, что стереохимическим следствием строения кобаламина оказывается его способность давать 5-координационные комплексы. Такая геометрия молекулы идеальна для быстрых каталитических реакций.[ ...]
До настоящего времени возможность участия координационных изомеров в биохимических превращениях не рассматривалась. Это, однако, не означает, что подобного рода изомёризационные (процессы не могут иметь места при биокаталитическом акте. Возможность образования сг-Со-С-связи в «нижнем» аксиальном положении, которая до сих пор не учитывалась, допускает новые предположения о характере контакта кофермента с субстратом.[ ...]
В обоих случаях энергия, необходимая для жизнедеятельности клеток, получается за счет окислительно-восстановительных реакций, суть которых состоит в переносе водорода с восстанавливающих молекул на окислительные. Часть выделяющейся при этом энергии запасается впрок в виде связей органических фосфатов типа АТФ или тиоэфирных связей КоА-произ-водных. Важнейшая, или "ключевая", особенность окислительно-восстановительных реакций состоит в цикличности восстановления и окисления кофакторов, т. е. восстановленные молекулы кофактора вновь окисляются и т. д. Основная биологическая функция этих коферментов — перенос водорода (Н+) и электронов (е ) от окисляющихся молекул пищевых субстратов. Являясь составными частями многих ферментных систем, никотинамидные коферменты контролируют метаболизм клетки, определяют направленность обратимых окислительно-восстановительных процессов.[ ...]
В настоящее время еще ни для одного пиридоксалевого фермента полностью не установлена природа функциональных групп активного центра, их взаимное расположение, последовательность аминокислотных остатков важнейших участков белка и целый ряд других данных по структуре фермента. В связи с этим естественно думать, что реальный механизм ферментативного трансаминирования является гораздо более сложным. Совокупность свойств белкового компонента и кофермента определяет возможность усиления на 6—7 порядков каталитической активности, заложенную в самом пиридоксальфосфате.[ ...]
Общая схема цикла представлена на рисунке 52. В цикл вступает активный ацетат, или ацетил-КоА. Сущность реакций, входящих в цикл, состоит в том, что ацетил-КоА конденсируется с щавелевоуксусной кислотой (ЩУК). Далее превращение идет через ряд ди- и трикарбоновых органических кислот. В результате ЩУК регенерирует в прежнем виде. В процессе цикла присоединяются три молекулы ШО, выделяются две молекулы СОа.и четыре пары водорода, которые восстанавливают соответствующие коферменты (ФАД и НАД).[ ...]
В вопросе о конформации НАД и НАДН следует выделить две стороны (в порядке важности и сложности): 1) конформация НАД в растворе и 2) конформация НАД р фермент-коферментном комплексе. Строгая ориентация в пространстве отдельных частей молекулы динуклеотида необходима для вполне определенного расположения нико- инамидного кольца относительно функциональных групп фермента, а также молекулы субстрата. Малейшее изменение углов валентных связей или межатомных расстояний в молекуле кофермента должно приводить к нарушению уникальной согласованности и эффективности каталитического процесса. Вследствие комплементарное™ молекулы кофермента участку связывания поверхности фермента (ко-фермент является «слепком» с участка связывания) изучение структуры НАД в фермент-коферментном комплексе дает непосредственную информацию о пространственном расположении функциональных групп активного центра фермента, взаимодействующих с коферментом.[ ...]
Электрон передвигается по направлению к НАДФ (О/В потенциал— 0,32) через ряд переносчиков, расположенных в порядке возрастания О/В потенциала. Электроны спонтанно текут в сторону более высоких окислительно-восстановительных потенциалов. Переносчик, воспринимающий электрон от П700, имеет Е0 — 0,55. Следующим переносчиком является железосодержащий белок ферредоксвн .(¿о — 0,43). От феррбдоксина электрон переносится на НАДФ (Е0—0,32).[ ...]
Эго важнейший этап гликолиза. Сущность его заключается в окислении альдегидной группы в карбоксильную. Окисление идет с выделением энергии. Сопряжеппо с этим идет образование 1,3-дифосфоглицери-повой кислоты (процесс, требующий затраты энергии). Таким образом за счет энергии окисления при участии неорганического -фосфата (Н3РО4 образуется макроэргическая фосфатная связь. Одновременно происходит восстановление кофермента НАД. Молекула этого фермента состоит из четырех идентичных субъединиц. Каждая субъединица представляет одиночную полипептидпуто цепь приблизительно из 220 аминокислотных остатков. Фермент содержит БН-группы, которые играют важную роль в окислении ФГА. Кофермеит НАД взаимосвязан с ферментом на всем протяжении процесса.[ ...]
Живая материя (живые организмы) состоит из множества органических соединений, которые представляют собой соединения углерода в виде крупных молекул (биологических макромолекул). Будучи просты по строению, биологические макромолекулы являются строительными блоками, из которых построены все современные организмы. Специфика организмов любого вида определяется набором присущих только им белков, которые в сочетании с нуклеиновыми кислотами в виде нуклеопротеидов выполняют роль субстрата жизни. Более простые молекулы, из которых состоят макромолекулы, выполняют в клетках несколько функций. Например, аминокислоты служат не только строительными блоками белков, но и являются предшественниками многих гормонов животных, алкалоидов и пигментов растений, а также других макромолекул, тогда как нуклеотиды, являющиеся строительными блоками нуклеиновых кислот, могут служить коферментами и переносчиками энергии.[ ...]
Биохимические реакции, составляющие основу обмена веществ и энергии, протекают в присутствии биологических катализаторов — ферментов (энзимов). Это соединения белковой природы, повышающие скорость химических реакций в живом организме. Ферменты катализируют превращения веществ, поступающих из окружающей среды и образующихся внутри организма. Биологические катализаторы, выделяемые во внешнюю среду и переводящие сложные соединения в форму, способную усваиваться организмом, называются экзоферментами. Биохимические реакции, протекающие внутри клетки, осуществляются при участии эндоферментов. По химическому строению ферменты могут быть простыми (протеины) или сложными (протеиды) белками. Активность ферментов-протеинов обусловлена наличием активных (каталитических) центров в белковой молекуле. Молекула ферментов-протеи-дов включает активные группы, содержащие сложные органические соединения или металлы (железо, кобальт и др.). Белковая часть молекулы фермента называется фероном (носитель), а добавочная— агоном (активная группа). Если агон может существовать самостоятельно, то он называется коферментом. Коферменты обычно являются промежуточными переносчиками электронов или некоторых атомов и атомных групп (водорода, аминогрупп и др.) от одних соединений к другим. В состав коферментов часто входят витамины.[ ...]
Успехи, достигнутые в области рентгеноструктурного анализа, кинетики переходных процессов и химического катализа за последние 20 лет, в корне изменили наши представления о ферментативном катализе и механизме действия ферментов. Данная монография представляет собой краткий обзор последних достижений в этой сфере и адресована студентам и аспирантам, уже прослушавшим соответствующие курсы по химии и биохимии. В книге в теоретическом и методологическом аспектах рассматриваются два вопроса: природа взаимодействия между ферментом и его субстратами, обусловливающего ферментативный катализ и специфичность действия фермента, и взаимосвязь между структурой фермента и механизмом ферментативного процесса. Обсуждаются экспериментальные подходы, позволяющие проводить прямые исследования ферментов на молекулярном уровне. Большое внимание уделяется, например, исследованию ферментативных реакций в предстационарных условиях, когда ферменты используются в концентрациях, сопоставимых с концентрациями субстратов, и можно непосредственно наблюдать за промежуточными фермент-содержащими соединениями. Кратко освещены проблемы взаимодействия ферментов с несколькими субстратами в стационарных условиях, а также некоторые вопросы химии коферментов и кофакторов.[ ...]