Такие рыбы, как карпы, плотва, сельди и другие, собираются в стаи ж перемещаются от одного места к другому. Стаи возникают не на все .время жизни, а только на некоторое время, большей частью в период миграции. У других рыб (обычно у хищников) стаи не образуются и ры->бы живут поодиночке или держатся очень маленькими группами. В связи с этим возник вопрос, одинаков ли обмен веществ у рыб, находящихся в стае и у одиночек.[ ...]
Мы приведем примеры с питанием сомиков и золотой рыбки (стр. 270), из которых будет видно, что групповое содержание рыб оказывает влияние и на их питание. Причем это влияние разное и зависит •от вида рыб: хищники они (сомики) или мирные (золотые рыбки).[ ...]
Шуэт (БсЬиеи, 1933, 1934) первый отметил, что золотая рыбка в .группе потребляет кислорода меньше чем одиночка. Алле и Остинг (в 1935 г.), проверяя эти работы, пришли к выводу, что здесь дело не в групповом эффекте, а в чисто методической погрешности. Рыбы, находящиеся в группе, выделяют больше азотистых веществ, чем находящиеся поодиночке. Выделяющиеся азотистые вещества оказывают влияние на химические реакции при определении кислорода по способу Винклера. Если пробу воды до фиксации кислорода предварительно обработать перманганатом, то эта методическая погрешность будет исправлена, и тогда получается одинаковый результат по дыханию рыб, находящихся в группе и поодиночке.[ ...]
Однако как бы ни были убедительны доказательства Алле и Остин-га, все же, как показали позднейшие опыты, групповой эффект существует.[ ...]
Шлайфер (ЭЫаНег, 1938, 1939) весьма остроумными опытами показал, что рыбки, находясь в группе, потребляют кислорода меньше, чем ■если они находятся поодиночке. Чтобы исключить химическое влияние .других экземпляров, Шлайфер помещал золотых рыбок поодиночке в аквариум с зеркальными стенками. Рыбка, сидящая в таком аквариуме, видела собственное изображение и вела себя так, как будто она находится в группе рыб. Одновременно Шлайфером было установлено, что рыбы в группе совершают меньше движений. В связи с разным количеством движений у рыб получается и разное потребление кислорода.[ ...]
Из исследований Шлайфера вытекает, что в основе своей групповой эффект сводится к разному количеству движений у рыб.[ ...]
М. Штефан в своей кандидатской диссертации показывает, что у разных рыб групповой эффект выражен в разной степени. У колюшки и тольяна (стайный образ жизни) он выражен сильнее, чем у зеркального карпа и карася, а у камбалы и маслюка (одиночный образ жизни) его совсем нет. Гольян в группе потребляет кислорода на 16,2% меньше, чем когда он находится поодиночке (средние данные), а колюшка соответственно на 15,6®/о меньше. У маслюка разницы В интенсивности потребления кислорода, когда он находится в группе и поодиночке, нет, а зеркальный карп и карась занимают среднее положение. У них потребление кислорода на 7—10% (средние данные) меньше, когда они находятся в группе, чем когда они находятся поодиночке. Количество движений, определенное у колюшки и маслюка в течение 29—30 часов, практически одинаковое, не зависящее от того, находятся рыбы в группе или поодиночке. Эти данные Штефан стоят в противоречии с данными Шлай-•фера, Фрая и других, которые считают это небольшое различие в интенсивности потребления кислорода как результат разницы в количестве движений у рыб, находящихся в группе и поодиночке.[ ...]
Гейер и Манн провели опыты с окунем несколько в ином методическом плане. Они употребляли проточную воду, соединяли последовательно дыхательные камеры, в которых дышали рыбы, и определяли дыха-«ие отдельно в каждой дыхательной камере. Хотя рыбы находились отдельно и не могли видеть друг друга, а также свое изображение, все же они потребляли кислород в разном количестве. Это указывает на сложность «группового эффекта». Рыба, находясь вместе с другими рыбами, воспринимает их не только органами зрения, но и рядом других каких-то рецепторов.[ ...]
Приведенные данные показывают, что вопрос о влиянии пребывания рыбы в группе на интенсивность ее дыхания далеко не решен.[ ...]
Наиболее могущественным фактором внешней среды, влияющим на обмен, является температура. Давно известно, что с понижением температуры рыбы становятся вялыми, малоподвижными. При определенной температуре (как нижней, так и верхней) рыбы прекращают движение и впадают в оцепенелое состояние, а иногда, даже погибают.[ ...]
Рисунки к данной главе:
| Потребление 02 рыбами при разной температуре |
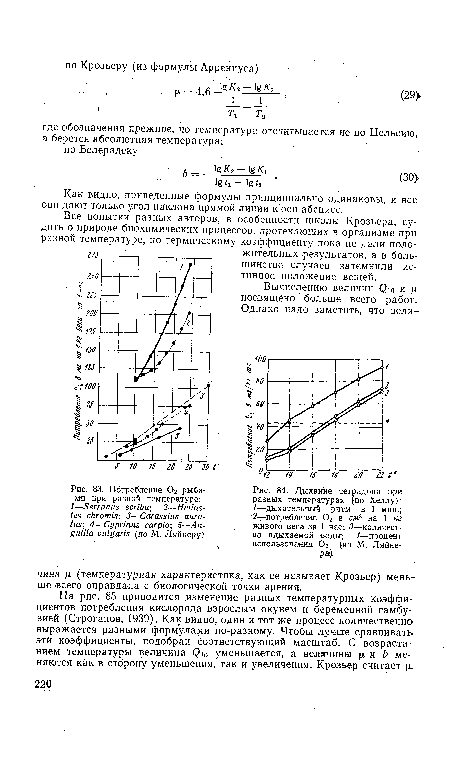 |
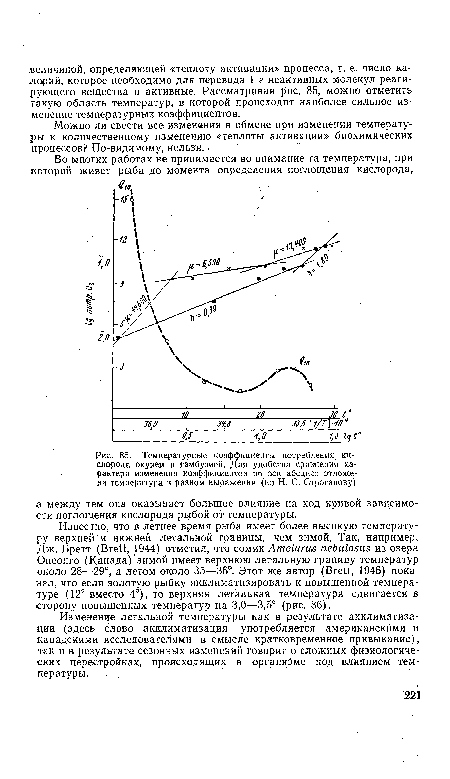
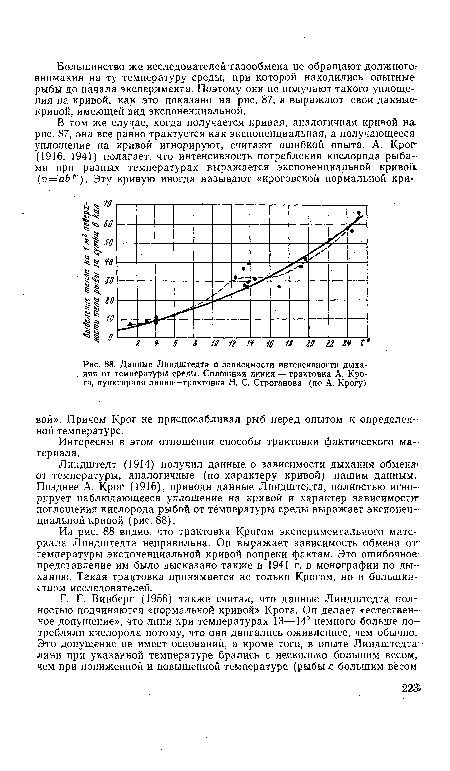
| Типичные кривые летальных температур для золотой рыбки, акклиматизированной к 4° (/) и к 12° (2)I (по Д. Р. Бретту) |
 |
| Интенсивность дыхания молодых гамбузий в зависимости от температуры среды. Сплошная линия—потребление СЬ, а пунктирная—выделение СО2. |
|
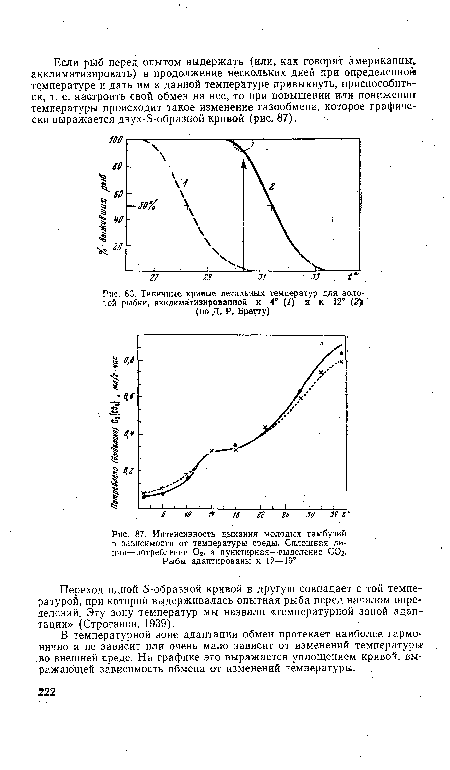
| Данные Линдштедта о зависимости интенсивности дыхания от температуры среды. Сплошная линия — трактовка А. Кро-га, пунктирная линия—трактовка Н. С. Строганова (по А. Крогу) |
 |
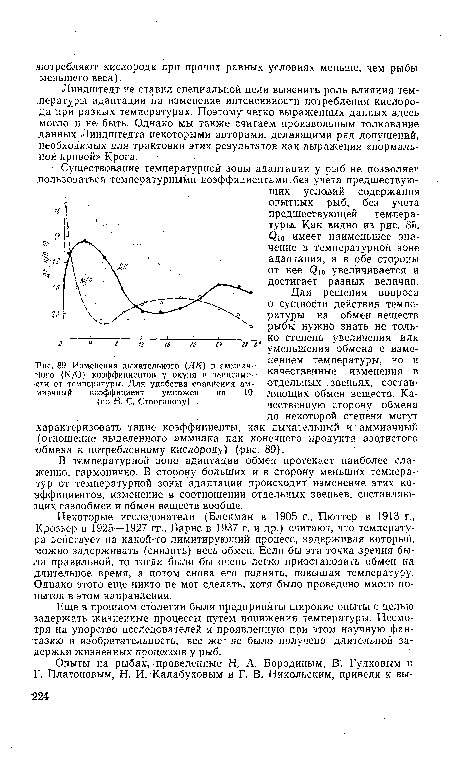
| Изменения дыхательного (ДК) и аммиачного (N/0) коэффициентов у окуня в зависимости от температуры. Для удобства сравнения аммиачный коэффициент умножен на 10 (по Н. С. Строганову) |
 |
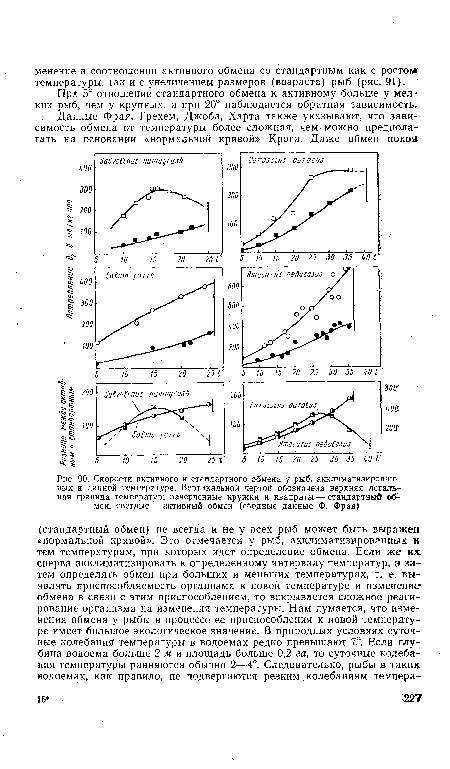
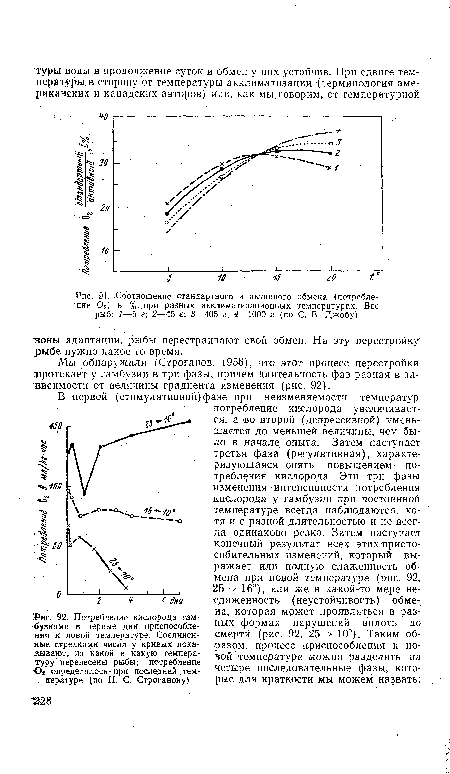
| Соотношение стандартного и активного обмена (потребление 02) в % при разных акклиматизационных температурах. Вес рыб |
 |
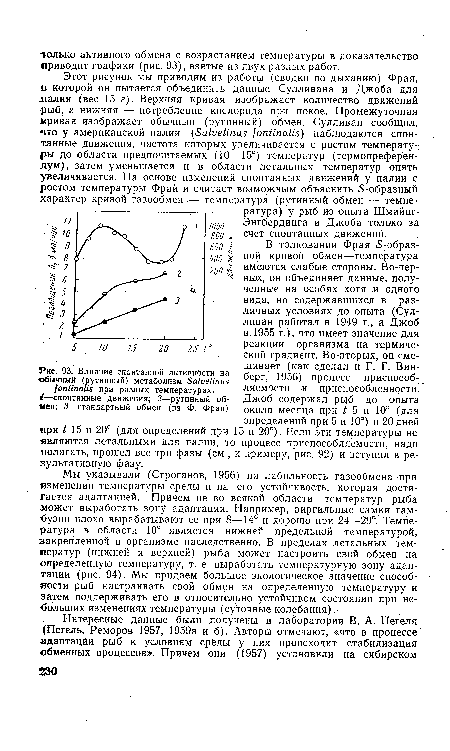
| Влияние спонтанной активности на обычный (рутинный) метаболизм БаЬеНпиз (опИпаЦй при разных температурах. |
 |