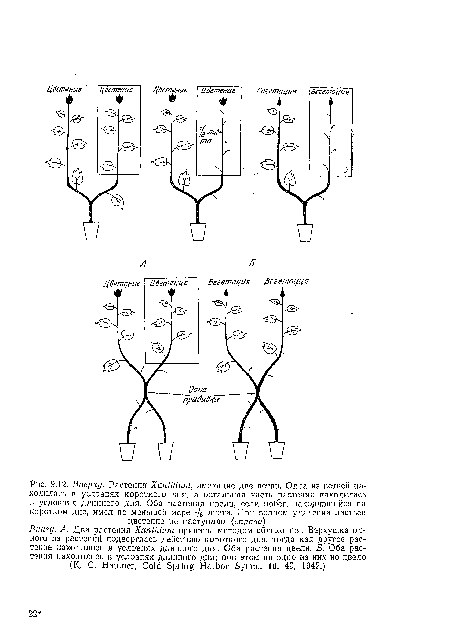
Еще более поразительный эксперимент был проведен на различных видах, в том числе на Xanthium и Perilla. С раетений-доиоров, находившихся на КД, удаляли по одному листу и прививали лист на растения, содержащиеся в условиях длинного дня. В положенное время растения-рецепторы зацветали, свидетельствуя о том, что стимул может передаваться из единственного листа.[ ...]
На длиннодневных растениях было выполнено значительно меньше прививочных экспериментов, чем на КДР, но полученные результаты, в сущности, схожи.[ ...]
Были сделаны попытки измерить скорость транспорта гормона. Один из методов заключался в использовании растений РкагЬШБ с раздвоенными побегами, у которых единственный лист-донор на одном из побегов подвергался действию условий КД; затем учитывалось время начала цветения второго побега (находящегося на ДД) в зависимости от разных расстояний между листом-донором и почкой-рецептором. Согласно данным, полученным такими, по-видимому, весьма косвенными методами, можно полагать, что скорость движения гормона намного меньше (2—4 мм/ч), чем обычная скорость перемещения сахаров по флоэме. Однако в экспериментах на РкагЬШБ были получены другие данные, свидетельствующие о более быстром транспорте (рис. 9.13). Согласно этим данным, стимул цветения переместился на расстояние 102 см за 2 ч, т. е. со скоростью 51 см/ч, что соизмеримо со скоростью движения ассимилятов по флоэме.[ ...]
Рисунки к данной главе:
Вернуться к оглавлению