Рассмотрим для начала какой-либо отдельный градиент Внешней среды, которым может быть, например, достаточно протяженный, ровный и непрерывающийся склон горы. Этот склон занят сообществами, в состав которых входит много видов растений, животных и сапробов. Мы уже знаем, что эти виды эволюционировали совместно, что их популяции связаны взаимными влияниями, что между некоторыми из них существует конкуренция и что их эволюция протекала в направлении ее снижения через дифференциацию экологических ниш. Какой же вариант распределения видов вдоль градиента является результатом этой эволюции? Ранее экологи предполагали, что виды образуют группировки, которые характеризуют определенные четко отграниченные друг от друга типы сообществ, часто называвшиеся ассоциациями. Такая точка зрения, когда сообщества признаются за отчетливо различимые единицы, может быть названа теорией сообществ-единиц. Она противоположна другому подходу, который независимо был развит в России в 1924 г. Л. Г. Раменским и в США в 1926 г. Г. А. Глизоном и известен как индивидуалистическая гипотеза. В основе этой гипотезы лежит представление о том, что виды распределены «индивидуалистически» и не образуют группировок, какие характерны для хорошо отграниченных друг от друга типов сообществ. Прежде чем отдать предпочтение одной из этих точек зрения (теории сообществ-единиц или индивидуалистической гипотезе), мы можем предложить несколько рабочих гипотез о возможных вариантах распределения видовых популяций, с тем чтобы после их обсуждения вновь вернуться к наблюдениям в природе и принять окончательное решение. Виды могут быть распределены по отношению друг к другу и к сообществам следующим образом.[ ...]
Изучением того, как виды в природе распределены по градиентам внешней среды, независимо друг от друга занимались автор, проводивший исследование в горах Грейт-Смоки-Маунтинс в 1947—1948 гг., и Дж. Кертис, работавший со своими студентами в штате Висконсин (первые результаты их наблюдений были опубликованы в 1951 г.). Подобного рода исследования могли быть осуществлены в горах, о которых шла речь в начале главы. Выборочные описания растительности с соответствующими измерениями значимости видовых популяций размещаются по высотному градиенту в направлении от вершин гор вниз к их подножию. Для эффективного изучения распределения популяций необходимо: 1) получить описания сравнимых местообитаний,например открытых склонов южной экспозиции на всех высотных уровнях, и 2) провести достаточно большое число описаний, с тем чтобы можно было сгруппировать их и вывести средине по высотным интервалам. Необходимо усреднить пять или более описаний на каждом 100-метровом интервале изменения высоты над уровнем моря, для того чтобы устранить имеющиеся отклонения в распределении популяции. В результате будет получена трансекта с таблицами, в которых показано, как каждый вид распределен в отношении к высотному градиенту, к другим видам и типам сообществ, которые можно было наблюдать. Такие же описания мы можем выполнить и по отношению к топографическому градиенту увлажнения. Этот градиент, определяемый влажностью почвы и состоянием атмосферы, простирается от влажных днищ каньонов через более сухие иижние части стенок каньонов, открытые северные склоны и промежуточные восточные и западные склоны вплоть до наиболее сухих южных и юго-западных склонов. Чтобы с помощью градиента увлажнения можно было получить топографическую трансекту, необходимо провести учеты на одной и той же высоте над уровнем моря. Из описаний, выполненных для равных либо очень сходных высот, выводятся средние для различных условий среды вдоль градиента увлажнения.[ ...]
Интересно выяснить, почему виды не эволюционируют в направлении образования групп с параллельным распределением. На рис. 3-4 мы уже показывали, как два вида, конкурирующие в отношении градиентов экологических ниш, имеют тенденцию дивергировать; отбор увеличивает различие между средними адаптивными позициями популяций двух видов по градиенту ниши. Заменим теперь градиент ниши (горизонтальная ось на рис. 3-4) на градиент местообитания. Два вида (в той же самой нише или в сходных) находятся в состоянии острой конкуренции в границах одного и того же значения какого-либо градиента местообитания, например высоты над уровнем моря или увлажнения. В этом случае отбор также будет увеличивать различия между средними положениями в адаптации видов вдоль градиента местообитания. Подобно тому кяк конкурирующие виды эволюционируют в направлении дифференциации экологических ниш, они эволюционируют также и р направлении дифференциации различий в местообитании.[ ...]
Приведем упрощенный пример этого процесса. Несколько видов распределены по градиенту условий среды (рис. 4-3), Новый вид, обозначенный цифрой 4, внедряемся на эту территорию. Его потенциальное распределение вдоль градиента, сформировавшееся в соответствии с генетическими свойствами вида в другом районе, показано пунктирной линией в верхней части рисунка. Однако поскольку вид 4 сказывается в состоянии жесткой конкуренции с видом 3, его популяция смещает свое положение в направлении к виду 5 и занимает некоторую часть градиента между этими двумя видами (3 и 5), что и отражено в иижней части рисунка. Это в свою очередь вызывает изменение в характере распределения других видов, отчего полоса их распределений, показанная отрезками прямых между верхним и нижним рисунками, уменьшается. Вид 4 удерживается и получает свое собственное место на шкале градиента благодаря кгк различию его ниши от ниш видов 3 и 5, достаточному, чтобы он мог существовать в их присутствии, так и благодаря новой амплитуде предпочитаемого местообитания, что снижает конкуренцию, возникающую от частичного перекрытия ниш.[ ...]
Здесь Sa — число видов в описании A; Sb — число видов в описании В; Sab число видов в обоих описан них.[ ...]
Здесь ра — выраженная в десятых долях аиачимость данного вида в описании А; рь — выраженная в десятых долях значимость того же вида в описании В.[ ...]
Эти меры выражают экологическое расстояние, то ес!ъ степень несходства видового состава описаний, отражающую их положение вдоль градиентов условий среды или других факторов. Более высокое бета-разнообразие градиента сообщества, помещенного внизу рис. 4-2, проявляется в более низком значении процентного сходства описаний в крайних точках градиента (0,9% против 18% для сообщества, показанного в верхней части рисунка, что соответствует и большему экологическому расстоянию между этими крайними точками).[ ...]
Рисунки к данной главе:
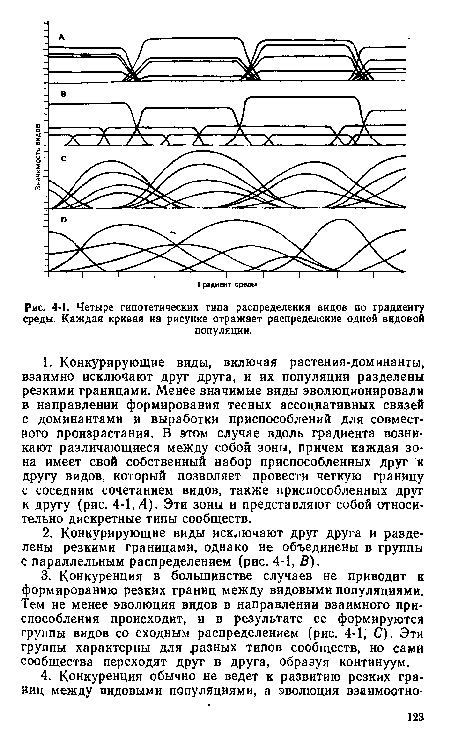
| Четыре гипотетических типа распределения видов по градиенту среды. Каждая кривая иа рисунке отражает распределение одной видовой |
 |