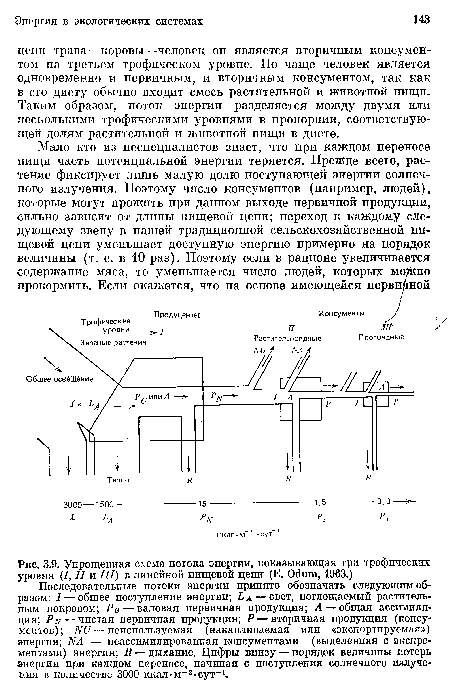
Принципы организации пищевых цепей и действия двух законов термодинамики можно уяснить, рассмотрев схемы переноса энергии на рис. 3.9—3.11. На этих схемах четырехугольники («резервуары») изображают трофические уровни, а линии («трубы») — потоки энергии от каждого уровня или к нему. Как требует первый закон термодинамики, приток энергии уравновешивается ее оттоком, и каждый перенос энергии сопровождается ее рассеиванием в форме недоступной для использования тепловой энергии (при дыхании), как того требует второй закон.[ ...]
На рис. 3.9 представлена сильно упрощенная схема потоков энергии на трех трофических уровнях. Здесь вводятся принятые в литературе обозначения разных потоков и показано, что на каждом последующем уровне поток энергии сильно уменьшается независимо от того, рассматриваем ли мы общий поток (/ и 4) или компоненты Р и /?. Показан и «двойной метаболизм» продуцентов (т. е. валовая и чистая продукция) и то, что на первом трофическом уровне поглощается около 50% падающего света а превращается в энергию пищи всего 1% поглощенной энергии (ср. с табл. 3.4). Вторичная продуктивность (Рг и Рз) на каждом последующем трофическом уровне консументов составляет около 10% предыдущей, хотя на уровне хищников, как показано на рис. 3.9, эффективность может быть выше, скажем 20%. Если питательная ценность источника энергии велика (например, продукт фотосинтеза, извлекаемый или выделяемый прямо из растительных тканей), то эффективность переноса энергии может быть гораздо выше. Но, поскольку и растения, и животные производят немало трудно перевариваемого органического вещества (целлюлоза, лигнин, хитин), а также химические ингибиторы, препятствующие поеданию возможными консументами, средняя эффективность переноса энергии между трофическими уровнями в целом составляет 20% и менее.[ ...]
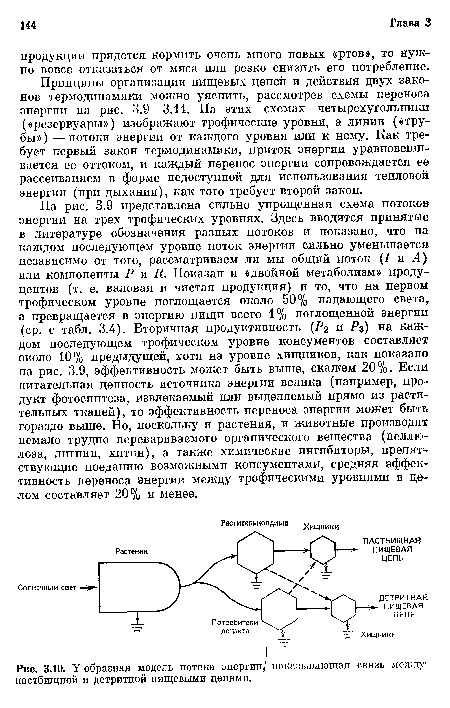
На рис. 3.10 пастбищные и детритные пищевые цепи показаны в виде Y-образной, или двухканальной, диаграммы потоков энергии. Эта модель более реалистична, чем простая одноканальная схема, поскольку 1) она соответствует основной ярусной структуре экосистем, 2) прямое потребление живых растений и использование мертвого органического вещества обычно разделены в пространстве и времени и 3) макроконсументы (фаготрофные животные) и микроконсументы (сапрофитные бактерии и грибы) сильно различаются отношениями интенсивности обмена к размерам организма и для их изучения требуются разные методы.[ ...]
Величины тех частей энергии чистой продукции, которые текут по двум путям, различны в экосистемах разного типа и часто варьируют по сезонам или по годам в одной и той же экосистеме. На некоторых мелководьях (например, в проливе Лонг-Айленд, изученном Райли, Riley, 1956) и на интенсивно используемых пастбищах и в степях по пастбищной цепи может идти 50 и более процентов чистой продукции. Напротив, марши, океаны, леса, да и большинство природных экосистем, функционируют как детритные системы; в них 90 и более процентов автотрофной продукции потребляется гетеротрофами только после того, как листья, стебли и другие части растений отмирают, подвергаются «переработке», превращаясь в диспергированное или растворенное органическое вещество, поступающее в воду, донные осадки и почву-Как подчеркивалось в гл. 2, такое отсроченное потребление увеличивает структурную сложность, а также накопительную и буферную емкость экосистем.[ ...]
В табл. 3.10 приведены данные, полученные при длительном исследовании влияния степени выпаса на естественное пастбище, проведенном на Великих равнинах (США). В первые несколько лет девятилетнего периода исследований выход мяса был максимален при очень интенсивном использовании пастбища, когда бычки изымали более 50% годовой чистой продукции, но через 9 лет пастбище оказалось настолько нарушенным, что выпас пришлось прекратить (т. е. выход упал до нуля). При низкой и умеренной интенсивности выпаса не только сохранялись выход мяса и хорошее состояние пастбища, но и «качество» поголовья было выше, о чем свидетельствовал больший вес одного теленка в конце сезона. Вопрос об оптимальном и максимальном потреблении еще будет обсуждаться в разделе о поддерживающей емкости среды.[ ...]
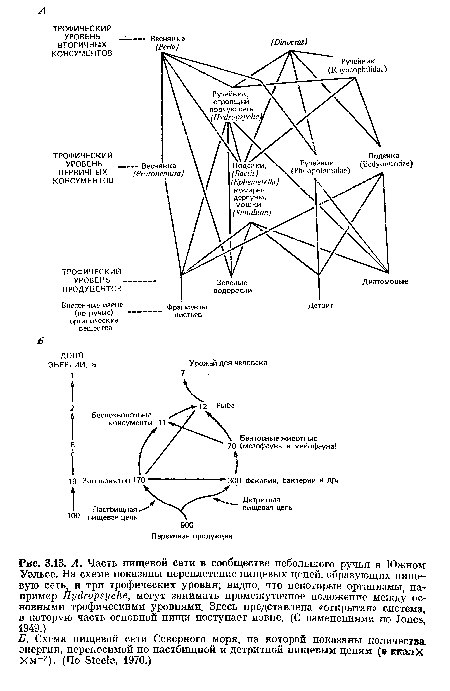
Как показано на рис. 3.11, А, две отдельные субсистемы пищевых цепей приурочены в основном к наземным или мелководным экосистемам; это зерновые пищевые цепи, начинающиеся с семян, которые представляют собой высококачественные источники энергии и составляют важную часть рациона людей и многих животных, и нектарные пищевые цепи, начинающиеся с нектара цветущих растений, которым для опыления требуются насекомые или другие животные. Сложные и мутуалистические взаимоотношения, развившиеся в эволюции между растениями и опылителями и растениями и зерноядными животными, обсуждаются в гл. 6.[ ...]
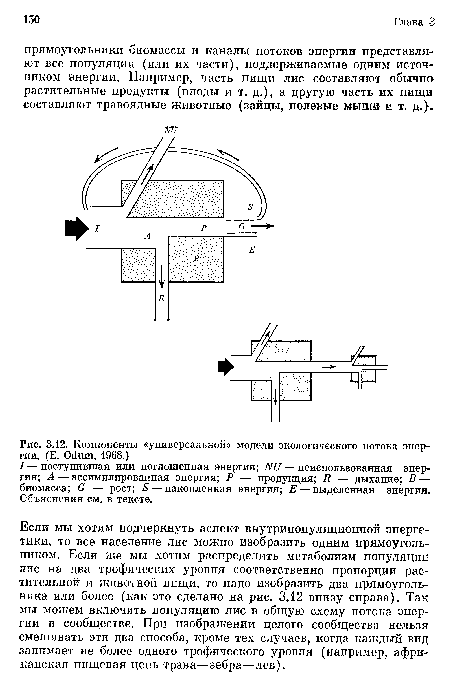
Универсальная модель потока энергии. Каковы основные компоненты модели потока энергии? На рис. 3.12 представлена модель, которую можно назвать «универсальной», поскольку она приложима к любому живому компоненту системы, будь то растение, животное, микроорганизм, особь, популяция или трофическая группа. Соединенные между собой, такие графические модели могут отразить, как уже показано, пищевые цепи или биоэнергетику экосистемы в целом. На рис. 3.12 квадрат, обозначенный буквой В, изображает живую структуру или биомассу основного компонента модели. Хотя биомассу выражают обычно через какую-либо массу (живая [сырая] масса, сухая масса или масса обеззоленного вещества), желательно выражать биомассу в калориях, чтобы выявить связь между размерами потока энергии и одномоментной или средней биомассой. Общее поступление энергии на рис. 3.12 обозначено буквой /. Для облигатных автотрофов это свет, для облигатных гетеротрофов — органическая пища. Как говорилось в гл. 2, некоторые водоросли и бактерии могут использовать оба источника энергии, а многие из них нуждаются в определенном соотношении этих источников. Сходная ситуация наблюдается у лишайников и тех беспозвоночных животных, в которых содержатся водоросли-мутуалисты. В таких случаях канал притока на схеме потоков энергии можно разделить пропорционально использованию различных источников энергии или же можно разделить биомассу на отдельные резервуары, если вы хотите, чтобы каждый резервуар соответствовал одному энергетическому (трофическому) уровню.[ ...]
Если мы хотим подчеркнуть аспект внутрипопуляционной энергетики, то все население лис можно изобразить одним прямоугольником. Если же мы хотим распределить метаболизм популяции лис на два трофических уровня соответственно пропорции растительной и животной пищи, то надо изобразить два прямоугольника или более (как это сделано на рис. 3.12 внизу справа). Так мы можем включить популяцию лис в общую схему потока энергии в сообществе. При изображении целого сообщества нельзя смешивать эти два способа, кроме тех случаев, когда каждый вид занимает не более одного трофического уровня (например, африканская пищевая цепь трава—зебра—лев).[ ...]
Но достаточно о проблеме источников энергии. Не вся энергия, поступающая в биомассу, подвергается превращению. Часть ее может просто пройти через биологическую структуру; например, пища может пройти через пищеварительный тракт, не включаясь в метаболизм, а часть энергии света проходит через растительность не усваиваясь. Эта часть энергии обозначена через NU. Используемая, или ассимилируемая, часть на схеме обозначена буквой А. Отношение А и /, т. е. эффективность ассимиляции, широко варьирует. Оно может быть очень мало, как в случае усвоения света растениями или ассимиляции пищи у животных-детритофагов, или же очень велико, как в случае животных или бактерий, питающихся высококалорийной пищей, например сахарами и аминокислотами.[ ...]
Рисунки к данной главе:
| Упрощенная схема потока энергии, показывающая три трофических уровня (I, II и III) в линейной пищевой цепи (Е. Odum, 1963.) |
 |
| У-образиая модель потока энергии/ показывающая связь между пастбищной и детритной пищевыми цепями. |
 |
| Компоненты «универсальной» модели экологического потока энергии. (Е. Odum, 1968.) |
 |