С широко распространенными растениями связано много видов насекомых, поскольку разнообразны условия существования, однако некоторые данные указывают на самостоятельный «эффект площади» расселения.— Там, где орляк редок, связанное с ним сообщество насекомых относительно ненасыщенно. — Разнообразие архитектоники растений влияет на видовое богатство насекомых-фитофагов. — На биологически «удаленных» растениях обитает меньше видов.[ ...]
Существование особого «эффекта площади» в случае насе-комых-фитофагов в какой-то мере доказывается тем обстоятельством, что на ряде растений, обилие которых (т. е. их «площадь») за последнее время возросло внутри некого конкретного местообитания, видовое богатство фитофагов увеличилось, в то же время как па растениях, сокративших свое обилие, снизилось и количество видов насекомых (Strong et al„ 1984). Однако не исключено, что с точки зрения насекомых изменения в обилии кормовых растений привели и к изменениям в разнообразии местообитаний.[ ...]
На рис. 20.14 приведен график зависимости «число видов — площадь» для растительноядной энтомофауны орляка четырех рассмотренных регионов. «Площадь» соответствует территории, занятой в каждом случае орляком, по оценкам Лотона. Хотя были взяты всего четыре точки, полученная зависимость между числом видов и площадью оказалась в высшей степени значимой статистически. По всей вероятности, в Нью-Мексико и на Гавайских островах с орляком связано мало насекомых-фито-фагов, поскольку само растение встречается здесь относительно редко. А поскольку этот эффект проявляется при сравнении отдельных участков, то, скорее всего, важную роль играет именно редкость как таковая, а не разнообразие местообитаний.[ ...]
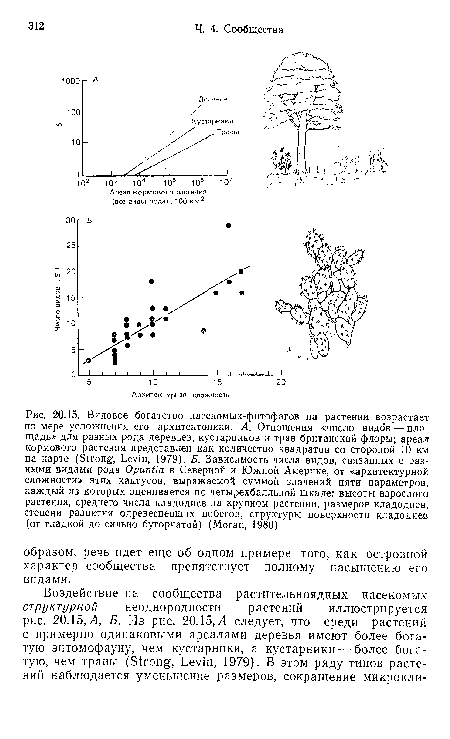
Совершенно очевидно, что архитектоника растения оказывает основное влияние на богатство сообществ растительноядных насекомых, но, как и в случае с островами, трудно решить, насколько это связано по отдельности с размерами (включая и число однородных частей) и с неоднородностью условий среды и ресурсов. По оценке Морана, у кактусов сами размеры обусловливают примерно 35% различий видового богатства, а все особенности архитектоники растения вместе взятые — 69%. Однако размерный диапазон у кактусов гораздо меньше, чем у деревьев и трав, сопоставленных на рис. 20.15, Л, так что различение эффектов величины и неоднородности в этих случаях требует дальнейших исследований.[ ...]
Влияние таксономической или биохимической «удаленности» на обеднение растений видами-фитофагами не всегда хорошо заметно при сравнении отдельных таксонов, однако ряд работ говорит о возможном существовании подобного эффекта. Например, в Калифорнии на двух древесных породах, не относящихся к дубам, но близких к роду Quercus, развивается меньше видов орехотворок, чем можно было бы ожидать исходя из данных по многочисленным здесь видам дубов (рис. 20.4.5; Cornell, Washburn, 1979). А среди британских однодольных меньше других насекомых-фитофагов имеют представители более бедных видами родов (рис. 20.16; Lawton, Schröder, 1977).[ ...]
Подведем итоги. Итак, больше видов растительноядных насекомых можно обнаружить на более крупных растениях с более сложной архитектоникой, на часто встречающихся и широко распространенных растениях и, вероятно, на растениях, представленных в одной области рядом близкородственных видов.[ ...]
Круговорот видов. — Обновление видов на островах приводит к неопределенности структуры сообщества.[ ...]
Положительная остаточная вариация.[ ...]
Равновесная теория Мак-Артура и Уилсона предполагает для острова не только наличие характерного видового богатства, но и постоянный круговорот видов, т. е. непрерывное поселение новых форм и вымирание уже присутствующих. Значит, конкретный состав островной биоты в каждый момент времени должен быть во многом случаен. Однако сам этот круговорот изучается редко, поскольку требует постоянного наблюдения за сообществами на протяжении длительного времени, что, как правило, технически сложно и дорого. Надежные данные по круговороту встречаются еще реже, так как предполагают всякий раз учет всех видов. Существенно и то, что любой полученный результат всегда дает заниженную оценку реального круговорота, поскольку наблюдатель не может в течение всей работы следить за всем происходящим одновременно.[ ...]
Экспериментальное подтверждение круговорота и неопределенности состава дано в работе Симберлоффа и Уилсона (Simberloff, Wilson, 1969), истреблявших всех беспозвоночных на нескольких мангровых островках архипелага Флорида-Кис и наблюдавших за их реколонизацией. Приблизительно через 200 дней видовое богатство стабилизировалось примерно на первоначальном уровне, но состав фауны существенно изменился. Эти данные позволили оценить скорость круговорота видов на островках как величину, составляющую 1,5 случая вымирания или иммиграции за год (Simberloff, 1976).[ ...]
Рисунки к данной главе:
Вернуться к оглавлению