Отсутствие сосуществования морфологически сходных видов на самом деле представляет собой лишь частный случай невозможности совместного обитания потенциальных конкурентов в целом. На ее основе Даймонд (Diamond, 1975), например, предложил действие конкуренции среди птиц архипелага Бисмарка. Однако Коннор и Симберлофф (Connor, Simberloff, 1979), использовав нейтральные модели, оспорили выводы, сделанные Даймондом и другими авторами на основе данных о распределении видов по островам. В построенных моделях сохранились всего три параметра реальной структуры сообществ: число видов на каждом острове (что отражало различия в размерах островов), число островов, заселенных каждым видом (это отражало различия в способностях видов к расселению и колонизации) и общий вид функции встречаемости каждого вида (см. разд. 18.3.3). В рамках этих ограничений Коннор и Симберлофф случайным способом составили сообщества из исходной совокупности видов (ее роль играла орнитофауна архипелага в целом). Они проанализировали данные по птицам островов Новые Гебриды (см. рис. 18.8), а также по птицам и летучим мышам Вест-Индии, во всех случаях придя к заключению, что реальные сообщества не имеют значимых отклонений от случайно подобранных.[ ...]
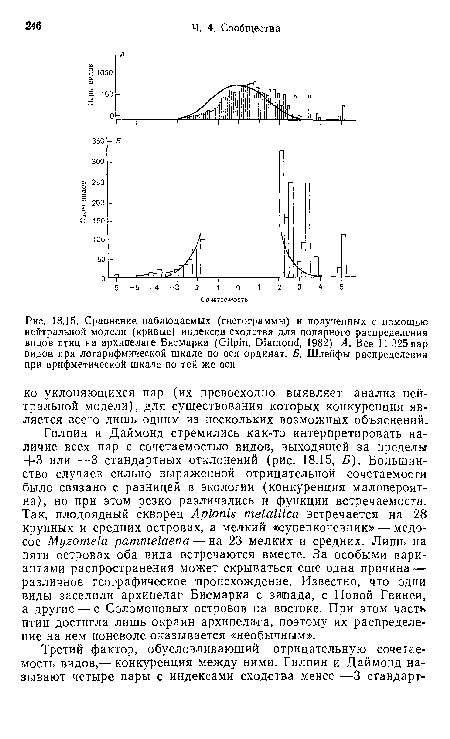
Если виды распределены так, как предсказывает случайная модель, индексы сходства будут распределены нормально со средним значением, равным нулю, и стандартным отклонением, равным единице. Однако для пар конкурирующих видов эти индексы будут заметно отрицательными (реальное число общих островов будет меньше ожидаемого).[ ...]
Положительная сочетаемость определялась такими факторами, как одинаковая функция встречаемости, общее географическое происхождение, эндемизм (распространение ряда видов ограничивалось всего одним из крупных островов архипелага), сходство местообитаний.[ ...]
В целом же, учитывая все сказанное выше, можно считать, что уточненный подход Гилпина и Даймонда к нейтральным моделям, раскрыл роль межвидовой конкуренции в организации сообществ птиц на архипелаге Бисмарка, хотя и в варианте этих авторов еще сохранялись многие недостатки первоначального анализа. Роль конкуренции была четко установлена лишь в немногих случаях, но предполагалась в гораздо большем их числе (там, где важное значение имеют функции встречаемости и возможна диффузная конкуренция). Столь же существенно по-ви-димому, следующее заключение Гилпина и Даймонда: даже если роль межвидовой конкуренции значительна, она все же остается лишь одним из нескольких основных факторов организации сообщества.[ ...]
Рисунки к данной главе:
Вернуться к оглавлению