Модель сукцессии Хорна (Horn, 1975, 1981) проливает некий свет на механизм смены видов. Этот автор признавал, что в гипотетическом лесном сообществе можно было бы предсказать изменения в составе древесных пород, зная две вещи: во-первых, для каждой породы — вероятность, с которой один экземпляр сменяется за определенное время экземпляром того же или иного вида; во-вторых, исходный видовой состав.[ ...]
Хорн полагал, что соотношение видов в лесном подросте отражает вероятность смены каждым из иих взрослого дерева, и оценил ее для срока 50 лет (табл. 16.3). Например, вероятность сохранения за березой желтой занимаемого ею на данный момент места — равна 5%, а смены ее ниссой лесной — 36%, кленом красным — 50 %, буком — 9%.[ ...]
Изучив распределение пологовых видов 25-летнего насаждения шт. Нью-Джерси, Хорн предсказал на основе своей модели изменения его состава на несколько сот лет вперед. Этот процесс в упрощенном виде представлен в табл. 16.4 для четырех пород.[ ...]
Ход этой гипотетической сукцессии позволяет выдвинуть ряд предположений. Так, клен красный должен быстро стать доми-нантом, а береза желтая обречена на исчезновение. Численность бука станет возрастать медленно, и он начнет доминировать позднее на фоне низкого обилия ниссы лесной и клена красного. Все эти предположения подтверждаются реальной картиной сукцессии.[ ...]
Наиболее интересный вывод из матричной модели Хорна — то, что по прошествии достаточного времени сообщество приобретает определенный стабильный состав, независимый от исходного. Конечный результат одинаков (он зависит исключительно от вероятностей замен), если начинать со 100% березы или бука (пока соседние районы обеспечивают приток семян исходно отсутствовавших видов), 50% ниссы и 50% клена или с любого другого сочетания.[ ...]
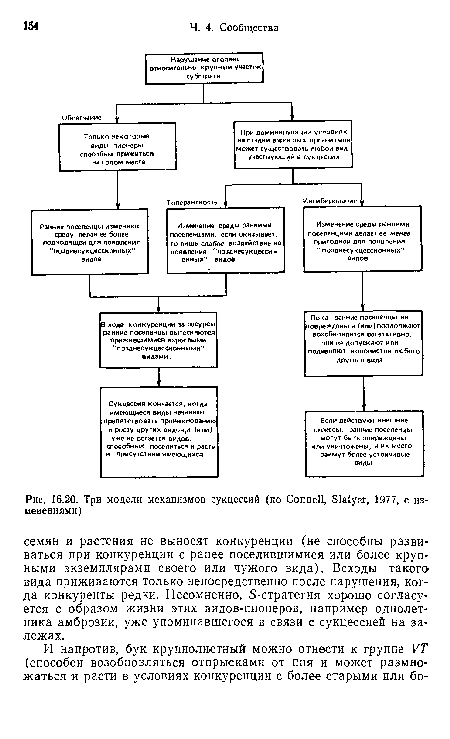
Примеры сукцессий в предыдущих разделах иллюстрируют различные, связанные с ними механизмы, включая облегчение и подавление (ингибирование), обзор которых дан в работе Коннелла и Слейтиера (Connell, Slatyer, 1977). Они предложили три модели, первая из которых (облегчения) — классическая, наиболее часто использовавшаяся в прошлом, в то время как две другие (толерантности и ингибирования), возможно, не менее важные, часто упускались из виду. На рис. 16.20 представлены основные черты всех трех.[ ...]
Отличительная особенность первой модели по сравнению с двумя другими — то, что развивающееся сообщество вызывает изменения абиотической среды, т. е. появление и рост определенных видов зависят от предыдущих, готовящих им условия. Наиболее яркие примеры связаны с первичными сукцессиями, например при отступлении ледника.[ ...]
Модель толерантности предполагает, что предсказуемая смена видов основана на их различных стратегиях потребления ресурсов. Виды более поздних стадий нуждаются в меньшем их количестве и способны достигать зрелости в присутствии более ранних видов, вытесняя их в конкурентной борьбе (пример — сукцессия на залежах).[ ...]
Модель ингибирования применяется при устойчивости всех видов к вторжению конкурентов, как показано выше для сукцессии макрофитов на валунах. Более поздние виды постепенно увеличивают свою численность, захватывая места, освобождающиеся при отмирании предшественников.[ ...]
Рисунки к данной главе:
| Три модели механизмов сукцессий (по Connell, Slatyer, 1977, с изменениями) |
 |