Сукцессии на заново осваиваемых территориях обычно идут несколько веков. Однако аналогичный процесс с участием мор-ских макрофитов, заселяющих недавно обнажившиеся участки скалистой литорали, занимает менее десяти лет. Жизни эколога вполне достаточно, чтобы проследить за всей сукцессией этих макрофитов, однако для непосредственного наблюдения за процессом отступления ледника она слишком мала. Правда, иногда можно собрать информацию и о более продолжительных событиях. Часто стадии сукцессии представлены градиентом сообществ в пространстве (пример — переход от соленого марша к лесу). Время появления того или иного сообщества можно установить, используя исторические карты, радиоуглеродный анализ и другие методики. Ход сукцессии могут отражать ряды сообществ, существующих в настоящее время, но соответствующих разным периодам времени, прошедшим с ее начала, однако вывод о том, действительно ли эти сообщества — разные стадии сукцессии, требует осторожности. Нужно помнить, например, что в умеренных северных широтах растительность местами еще ре-колонизирует пространство, освободившееся после последней ледниковой эпохи (гл. 1).[ ...]
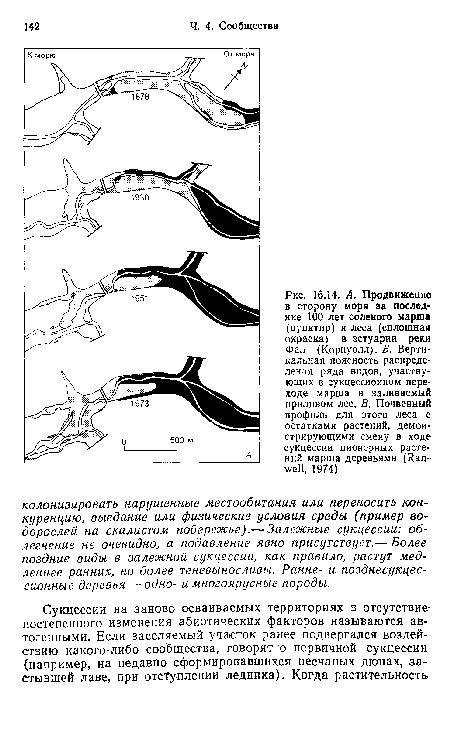
Первые виды в сукцессии могут настолько изменить условия или доступность ресурсов в местообитаниях, что становится возможным поселение новых видов. Этот процесс известен как облегчение (Connell, Slatyer, 1977). Он особенно важен при первичной сукцессии, когда исходные условия суровы, например на обнаженной почве после отступления ледника. Необыкновенно быстрое разледенение отмечено в районе бухты Глейшер (юго-восток Аляски). Примерно с 1750 г. ледник отступил здесь почти на 100 км, оставив морены, возраст которых может быть оценен по числу годичных колец наиболее старых, поселившихся на них деревьев. На последней моренной гряде максимальный возраст деревьев около 200 лет, а по мере приближения к леднику снижается. За последние 80 лет информация получена в ходе прямых наблюдений.[ ...]
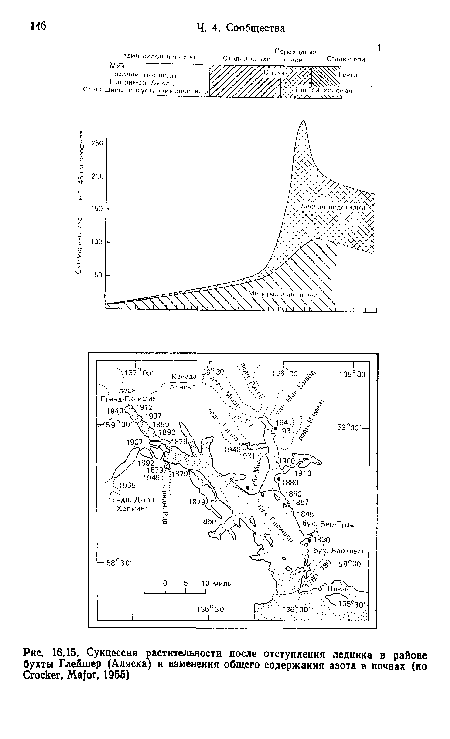
Одна из основных движущих сил сукцессии — изменение почвы первыми колонистами. И куропаточья трава, и ольха имеют симбионты, фиксирующие атмосферный азот (см. гл. 13), что приводит к накоплению в почве больших его запасов (рис. 16.15). Ольха также способствует сильному подкислению почвы, снижая за 50 лет pH ее поверхности приблизительно с 8,0 до 5,0. После этого ситхинская ель, используя накопленный азот, может поселяться и замещать ольху. Постепенное запасание в почве углерода ведет к развитию ее мелкокомковатой структуры, повышению аэрации и влагоемкости субстрата.[ ...]
На хуже дренированных склонах эта сукцессия не всегда заканчивается елово-тсуговым лесом. Во влажных местах, где уклона нет или он невелик, бурно развиваются сфагновые мхи, накапливающие воду и сильно подкисляющие почву, которая пропитывается влагой и испытывает дефицит кислорода; при этом большинство деревьев гибнет. Редко произрастающие сосны (Pinus contorta) — единственные деревья, способные выдержать плохую аэрацию субстрата возникающего болота. Следовательно, завершающая сукцессию растительность явно связана с локальными условиями.[ ...]
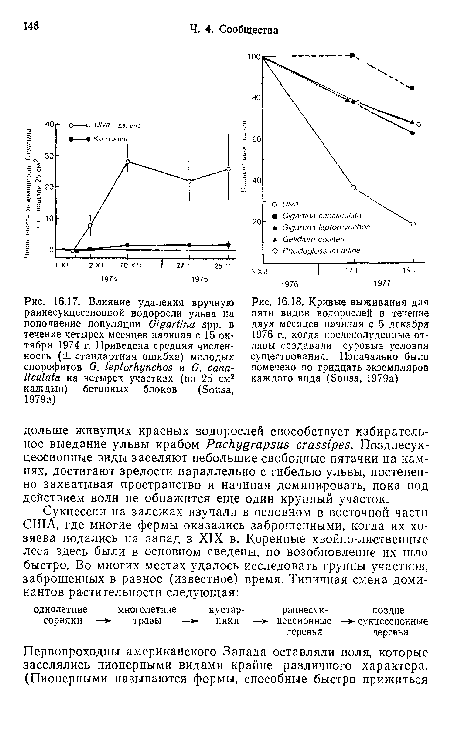
В южной части калифорнийского побережья водоросли разных видов достаточно закономерно сменяют друг друга на валунах нижней литорали (Sousa, 1979а). Основная естественная причина освобождения здесь пространства — перевертывание камней волнами. Водоросли реколонизируют свободные поверхности либо путем вегетативного роста сохранившихся экземпляров, либо, что бывает чаще, развиваясь из спор. Типичный ход естественной сукцессии на валунах можно моделировать либо искусственно очищая их, либо устанавливая на литорали бетонные блоки. Такой подход позволил точно описать сукцессию, а также выявить участвующие в ней механизмы.[ ...]
Озимые однолетники (главным образом другие сложноцветные) часто быстро вытесняют незимующие виды, в том числе и амброзию. У озимых трав мелкие семена, почти или вообще не имеющие стадии покоя, но расселяющиеся на большие расстояния. Прорастая вскоре после падения на землю (обычно в конце лета — осенью), они развиваются в зимующие розетки. Следующей весной они начинают вегетировать раньше незимующих однолетников, перехватывая у них таким образом ресурсы в виде света, воды, пространства и биогенных элементов. В противоположность пионерным однолетникам у растений более поздних стадий сукцессии, особенно в климаксовом лесу, семена не требуют света для прорастания, и если у первых под пологом деревьев семенное возобновление маловероятно, то у последних оно вполне возможно.[ ...]
Жизненный цикл раннесукцессионных растений короток. Их непрерывное существование зависит от переноса в другие нарушенные места. I эспособные выдержать конкуренцию с появляющимися позднее видами, они вынуждены быстро расти (основное их свойство), эффективно используя доступные ресурсы. В связи с этим скорость фотосинтеза на единицу площади листа (табл. 16.2) и относительная скорость роста (возможно, из-за накопления в растениях очень большого количества старой опорной ткани) с течением сукцессии в целом снижаются.[ ...]
Рисунки к данной главе:
| Сукцессия растительности после отступления ледника в районе бухты Глейшер (Аляска) и изменения общего содержания азота в почвах (но Crocker, Major, 1955) |
 |
| Кривые выживания для |
 |
| Теоретические кривые светового насыщения фотосинтеза для растений ранних, средних и заключительных стадий сукцессии (Ваггаг, 1979) |
 |