Между отдельными органами, тканями и даже клетками растения существуют сложные взаимодействия. Эти взаимодействия мы называем корреляциями (см. также гл. 1). Гармоничное развитие растения, т. е. согласованное развитие и рост отдельных его частей,— это результат коррелятивных взаимодействий. Поскольку растения обычно растут в течение всей жизни, самая большая роль принадлежит ростовым корреляциям. Различают коррелятивную стимуляцию и коррелятивное торможение роста. К первой категории относятся, например, ускорение роста в длину у колеоптилей злаков под влиянием верхушки колеоптиля, стимуляция деятельности камбия растущими почками, стимуляция роста корня побегом (рис. 14.1) и роста побега корнем, стимуляция роста плодов семяпочками или семенами, а также многие другие явления.[ ...]
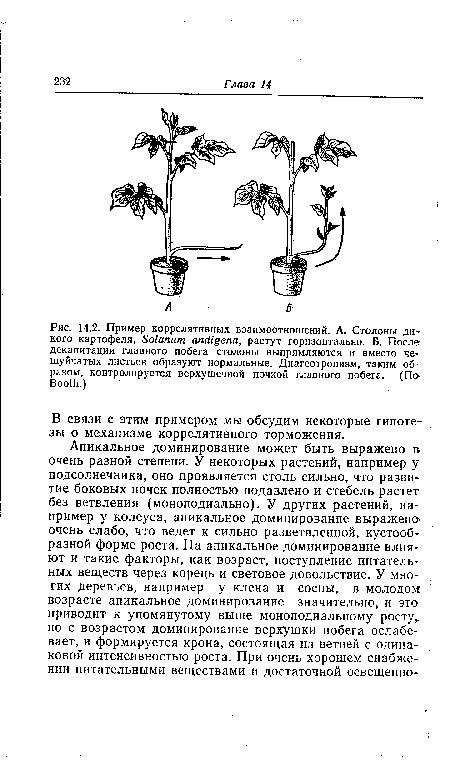
Коррелятивное тормозящее влияние оказывает, например, верхушечная почка на боковые почки, а кончик корня — на зачатки боковых корней. Упоминавшееся выше подавление опадения листовых черешков под влиянием листовой пластинки (разд. 12.2), так же как и торможение вегетативного роста растущими плодами, — это тоже примеры коррелятивного торможения. От коррелятивных воздействий зависит и направление роста боковых побегов. Побеги, растущие косо или даже горизонтально, нередко поднимаются, если удалить верхушечную почку (рис. 14.2). Важную роль играют коррелятивные взаимодействия также в дифференцировке и в изменении обмена веществ. Из многочисленных примеров мы упомянем лишь два: стимуляцию дифференцировки проводящих пучков под влиянием почек (наблюдаемую, в частности, в тканевых культурах) и стимулирующее влияние зародыша на синтез амилазы в зерне злаков.[ ...]
К наиболее изученным ростовым корреляциям относится торможение боковых почек растущей верхушечной почкой, называемое также апикальным доминированием.[ ...]
В связи с этим примером мы обсудим некоторые гипотезы о механизме коррелятивного торможения.[ ...]
Если удаляют верхушечную почку или же срезают весь растущий концевой побег, то заторможенные боковые почки трогаются в рост и образуют боковые побеги, в свою очередь конкурирующие между собой за доминантность и подавляющие друг друга.[ ...]
Листья, особенно молодые, также оказывают коррелятивное тормозящее влияние — прежде всего на боковые почки, которые могут быть в их пазухах. Поэтому для полного снятия коррелятивного торможения почек, например у побегов древесных растений, производят декапитацию и удаление листьев.[ ...]
Еще в начале изучения ауксинов было выяснено, что действие апикальной почки на боковые почки может быть заменено действием ауксиновой пасты (см. 2.5). Этот эффект, впервые отмеченный Тиманом и Скугом в 1934 г., с тех пор получил подтверждение в многочисленных экспериментах на растениях как с сильным, так и со слабо выраженным апикальным доминированием. Вряд ли можно сомневаться в том, что торможение боковых почек обусловлено ауксином, который образуется в верхушечной почке или молодых листьях и переходит отсюда в органы, расположенные ниже. Однако вопрос о том, как ауксин осуществляет это торможение, остается весьма спорным. Первая гипотеза, предложенная Тиманом, согласно которой ауксин верхушечной почки непосредственно тормозит боковые почки, не подтвердилась. Гипотеза Тимана была основана на двух предпосылках: 1) ауксин из главного побега передвигается в боковые почки, а это означает, что в них происходит акропеталь-ный транспорт; 2) боковые почки отвечают на воздействующий со стороны главного побега ауксин торможением роста, т. е. оптимальными для их роста должны быть более низкие концентрации ауксина. Обе предпосылки оказались неверными. Более того, выяснилось, что боковые почки содержат ауксин в субоптимальных концентрациях и при прямом нанесении ауксина их рост не только не затормаживается, а даже ускоряется.[ ...]
Согласно другой гипотезе, предложенной Сноу (Snow, 1937), существует корреляционный ингибитор, который образуется в коррелятивно заторможенных органах и, в отличие от ауксина, может проникать в боковые почки акропетально. В некоторых работах последних лет были сделаны попытки установить связь между этим гипотетическим гормоном и ингибиторами роста типа абсцизо-вой кислоты. Действительно, в коррелятивно заторможенных боковых почках, например у побегов клена и сирени, было обнаружено значительно больше абсцизо-вой кислоты, чем в тканях листа или главного побега.[ ...]
Однако после удаления листьев и декапитации содержание абсцизовой кислоты в расчете на одну почку не снижалось в той мере, как того следовало бы ожидать, если бы торможение было вызвано абсцизовой кислотой. Несмотря на это противоречие, гипотеза об участии абсцизовой кислоты в коррелятивном торможении почек весьма привлекательна. Мы уже упоминали, что покоящиеся почки многих растений отличаются повышенным содержанием абсцизовой кислоты (разд. 5.2.3.3). Коррелятивно заторможенные почки тоже можно рассматривать как покоящиеся (коррелятивный покой), и, как известно, у многих древесных растений коррелятивный покой может прямо переходить в состояние внутреннего зимнего покоя.[ ...]
Гиббереллины, по-видимому, не участвуют в гормональном контроле коррелятивного торможения почек. Гиббереллин не может заменить доминирующую верхушку, а воздействие гиббереллином на коррелятивно заторможенную почку не вызывает ее распускания. Однако обработка им интактной верхушечной почки усиливает апикальное доминирование. Это может быть обусловлено двумя причинами: индукцией роста, приводящей к усилению аттрагирующей активности, и повышением уровня ауксина в растении (см. разд. 3.2.1).[ ...]
Рисунки к данной главе:
| Пример корреляций |
 |