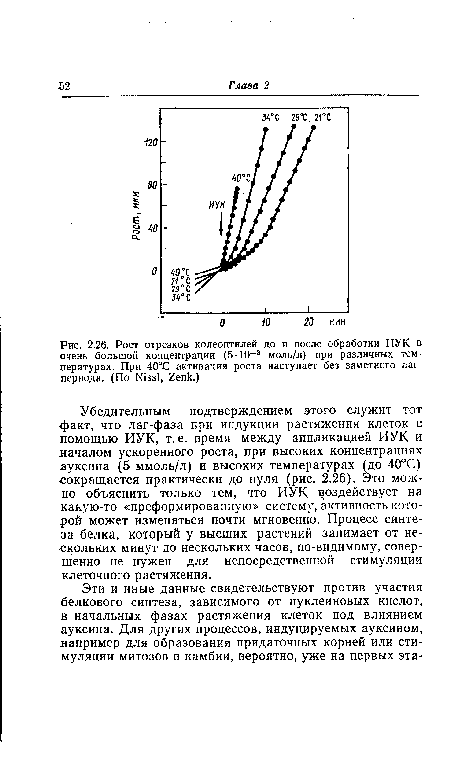
Убедительным подтверждением этого служит тот факт, что лаг-фаза при индукции растяжения клеток с помощью ИУК, т. е. время между аппликацией ИУК и началом ускоренного роста, при высоких концентрациях ауксина (5 ммоль/л) и высоких температурах (до 40°С) сокращается практически до нуля (рис. 2.26). Это можно объяснить только тем, что ИУК воздействует на какую-то «преформированную» систему, активность которой может изменяться почти мгновенно. Процесс синтеза белка, который у высших растений занимает от нескольких минут до нескольких часов, по-видимому, совершенно не нужен для непосредственной стимуляции клеточного растяжения.[ ...]
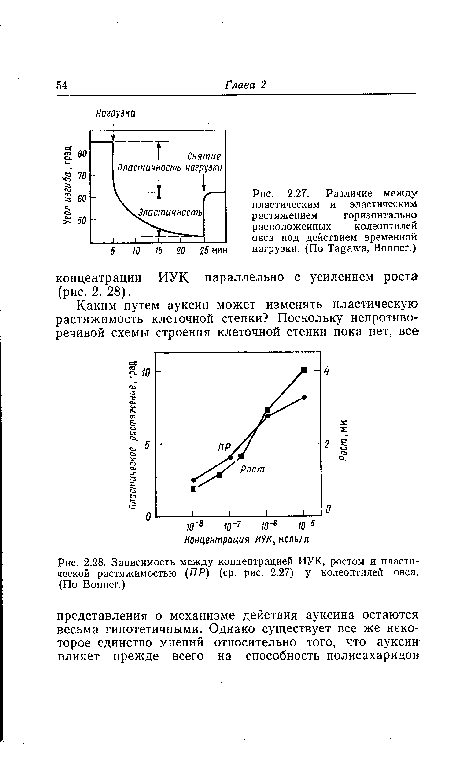
Растяжимость клеточной стенки можно подразделить на две составляющие: пластическую и эластическую растяжимость. Их можно различить с помощью несложных методических приемов (рис. 2.27): на верхушку горизонтально ориентированного колеоптиля помещают груз, вследствие чего она изгибается книзу. После снятия нагрузки верхушка колеоптиля за счет своей упругости поднимается вверх, не достигая, однако, исходного положения, поскольку одновременно она подверглась и пластической деформации.[ ...]
Увеличение растяжимости клеток под влиянием ауксинов пытались также объяснить усиленным метилированием пектинов, так как в результате этого процесса свободные карбоксильные группы замещаются метальными, что препятствует образованию кальциевых мостиков между молекулами пектиновых кислот. Однако и это представление оказалось несостоятельным, когда выяснилось, что индукцию роста ауксином не подавляют даже ингибиторы метилирования, например этионин.[ ...]
Наибольший интерес привлекает сейчас вопрос о том, каким образом ауксин изменяет способность белков стенки к ее разрыхлению. Маловероятно, чтобы ауксин действовал на эти ферменты непосредственно, например по-принципу аллостерического активирования. Более того, имеются многочисленные данные в пользу того, что одна из первых реакций клетки на ИУК состоит в активном выбросе протонов в клеточную стенку. Ионы Н+ могли бы в свою очередь активировать гидролитические ферменты клеточной стенки с низким оптимумом pH.[ ...]
Если согласно уравнению Лайнуивера—Берка построить график в координатах величин, обратных концентрации ИУК и скорости роста (для низких концентраций ИУК), то получится прямая линия (рис. 2.36); этот график позволяет вычислить константу Михаэлиса. Для взаимодействия ИУК с гипотетическим рецептором в коле-оптилях овса эта константа составляет около 6-10 8 моль/л. Таким образом, взаимоотношения между ИУК и растущей системой формально могут описываться так же, как взаимоотношения между ферментом и субстратом.[ ...]
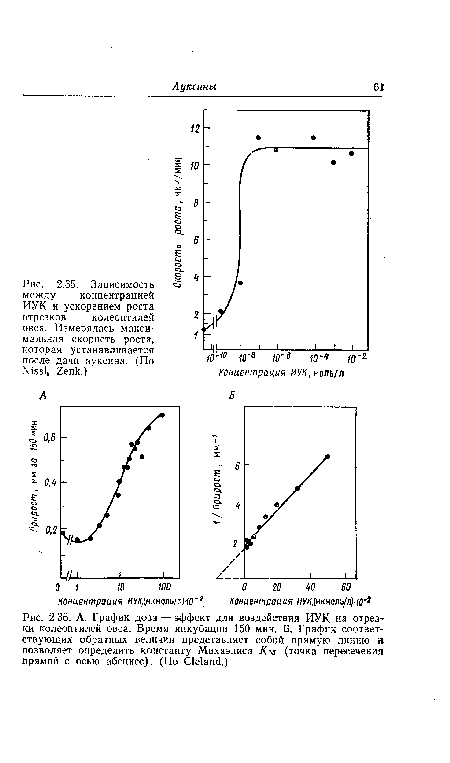
Для объяснения эффекта торможения был предложен ряд гипотез. Согласно теории «двухточечного взаимодействия» (см. разд. 2.4), молекулы ауксина при возрастающей конкуренции за место присоединения начинают все чаще связываться с рецептором не в двух точках, а только в одной. В результате этого биологическая активность молекулы не может проявиться. По-видимому, таким образом можно объяснить нисходящую часть графика доза—эффект у побегов. Однако представленная на рис. 2.35 сигмоидная кривая доза—эффект при коротких периодах воздействия ауксина говорит против теории двухточечного присоединения.[ ...]
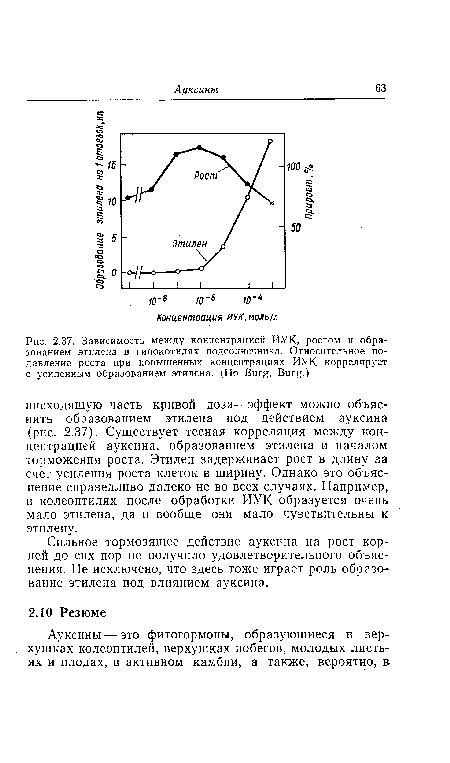
Сильное тормозящее действие ауксина на рост корней до сих пор не получило удовлетворительного объяснения. Не исключено, что здесь тоже играет роль образование этилена под влиянием ауксина.[ ...]
Рисунки к данной главе:
| Различие между пластическим и эластическим растяжением горизонтально расположенных колеоптилей овса под действием временной 5 10 15 20 25 мин нагрузки. (По Tagawa, Bonner.) |
 |
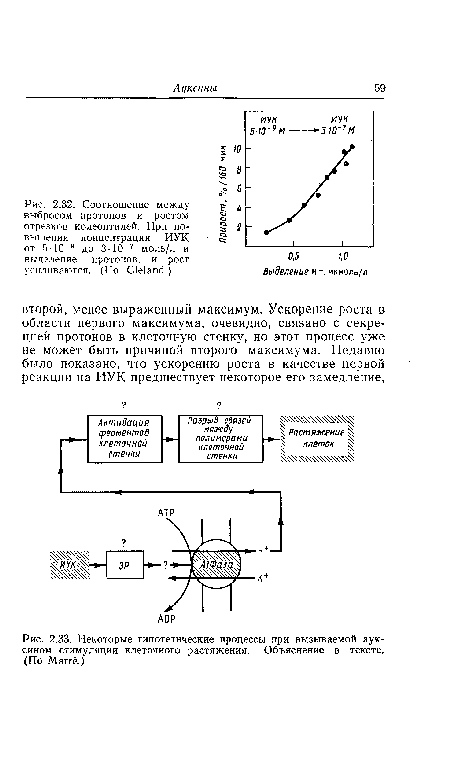
| Зависимость между концентрацией ИУК, ростом и пластической растяжимостью (ПР) (ср. рис. 2.27) у колеоптилей овса. (По Bonner.) |
|
| Соотношение между выбросом протонов и ростом отрезков колеоптилей. При повышении концентрации ИУК от 5-10 |
 |
| Некоторые гипотетические процессы при вызываемой ауксином стимуляции клеточного растяжения. Объяснение в тексте. (По Marré.) |
|
| Зависимость между концентрацией ИУК и ускорением роста отрезков колеоптилей овса. Измерялась максимальная скорость роста, которая устанавливается после дачи ауксина. (По N¡551» 2епк.) |
 |