Рассмотренные выше модели популяционного роста представляют собой «идеальные» схемы, основанные на видовьгх особенностях механизмов самоподдержания популяций и наиболее генеральных характеристиках среды обитания. Сложность условии, в которых разворачивается функционирование естественных видовых популяций, приводит к тому, что их состав, особенности структуры, обеспеченность пшцей и взаимоотношения с популяциями других видов никогда не остаются постоянными, демонстрируют количественные и качественные изменения во времени. Наиболее генерализованные из них проявляются в виде изменений численности популяций, которые могут быть относительно незакономерными, вызванными критическими воздействиями каких-либо факторов, но могут и иметь характер закономерных сезонных или (и) многолетних циклов. Последние привлекают особое внимание исследователей, поскольку циклические изменения численности отражают всю сумму реакций вида (его конкретных популяций) на сложный комплекс факторов абиотической среды, систему межвидовых отношений и изменения внутрипопуля-ционных взаимодействий.[ ...]
Вообще, появление в эволюции каких-либо форм заботы о потомстве четко коррелирует со снижением видовой нормы плодовитости: у таких видов существенно снижается смертность в раннем возрасте, а сохранение высокого репродуктивного потенциала при сниженной смертности биологически невыгодно. У видов, выкармливающих свое потомство, видовая норма плодовитости в большей степени зависит не от уровня смертности, а от возможности обеспечить выводок кормом (Д. Лэк, 1957).[ ...]
Обратная пропорциональность связывает плодовитость со средней продолжительностью жизни, свойственной данному виду: более долгоживущие виды отличаются меньшей плодовитостью (С.А Северцов, 1936; D. Promyslow, Р. Harwey, 1980). По суш, это тоже отражает корреляцию плодовитости и смертности, но в этом случае речь идет об уровне смертности, определяемом не внешними факторами, а физиологией вида.[ ...]
Видовые параметры плодовитости отражают, таким образом, средний уровень гибели, свойственный данному виду в многолетнем масштабе. В реальной обстановке конкретные величины как плодовитости, так и смертности испытывают постоянные колебания; соответственно этому меняется и уровень численности популяции и тенденции ее изменений во времени. На этой основе легко объяснить, а подчас и прогнозировать изменения численности, определяемые направленными сдвигами отдельных факторов и их комплексов.[ ...]
Типы динамики численности и экологические стратегии. Накопленный в настоящее время материал по разным группам живых организмов показывает, что численность естественных популяций не остается постоянной даже при выходе на плато логистической кривой. Более того, наряду с незакономерными и в большинстве случаев недолговременными изменениями численности, прямо связанными с положительным или отрицательным влиянием конкретных факторов, практически у всех исследованных видов обнаруживаются закономерные чередующиеся подъемы и спады численности, имеющие волнообразный, циклирующий характер и нередко охватывающие большие пространства. Такой характер динамики численности известен для насекомых (ГА Викторов, 1967), рыб (Г.В. Никольский, 1965), птиц (Д. Лэк, 1957), многих видов млекопитающих (СА. Северцов, 1936, 1941, 1942; И.Я. Поляков, 1949; А.А. Максимов, 1984) и других животных. Именно такого рода колебания обычно и имеют в виду, когда говорят о проблеме динамики численности.[ ...]
Лабильный тип динамики отличается закономерными колебаниями численности с периодами порядка 5—11 лет и более значительной амплитудой (численность меняется в десятки раз). Характерны сезонные изменения обилия, связанные с периодичностью размножения. Такой тип динамики характерен для животных разного, но, как правило, не крупного размера с более коротким сроком жизни (цо 10—15 лет) и соответственно более ранним половым созреванием и более высокой плодовитостью, чем у представителей первого типа. Повышена и средневидовая норма гибели. К этому типу динамики из млекопитающих относятся крупные грызуны, зайцеобразные, некоторые хищные; таков же общий характер динамики населения у многих птиц, рыб, насекомых с длинным циклом развития и некоторых других животных.[ ...]
Эфемерный тип динамики отличается резко неустойчивой численностью с глубокими депрессиями, сменяющимися вспышками «массового размножения», при которых численность возрастает подчас в сотни раз. Перепады ее от минимума до максимума осуществляются очень быстро (иногда в течение одного сезона); столь же. быстро происходит спад численности, который в таком случае часто называют «крахом популяции». Общая длина цикла обычно составляет до 4—5 лет, в течение которых «пик» численности занимает чаще всего не более одного года; у некоторых животных (например, у мелких грызунов) на эти короткие циклы «накладываются» более продолжительные (10—11 лет), но часто такие «большие волны» более выражены охваченным вспышкой пространством, чем уровнем численности. Резко выражены сезонные колебания обилия особей.[ ...]
Эфемерный тип динамики характерен для короткоживущих (не более 3 лет) видов с несовершенными механизмами индивидуальной адаптации и соответственно с высокой нормой гибели. Это некрупные животные, отличающиеся большой плодовитостью. Наиболее характерен такой тип динамики для мелких грызунов и многих видов насекомых с коротким циклом развития, но встречается и в других группах животного мира.[ ...]
Именно эта мысль лежит в основе концепции экологических стратегий, разработанной Р. Мак-Артуром и Э. Уилсоном (К. Мае АЛЬиг, Е. Vilson, 1967) и получившей широкое признание в современной экологии. Суть этой концепции сводится к тому, что успешное выживание и воспроизводство вида возможно либо путем совершенствования адаптированное™ организмов и их конкурентоспособное™, либо путем интенсификации размножения, что компенсирует повышенную гибель особей и в критических ситуациях позволяет быстро восстановить численность. Первый путь назван « -стратегией»; представители этого типа — чаще всего крупные формы с большой продолжительностью жизни; численность их лимитаруется преимущественно внешними факторами (коэффициент К в уравнении логистической модели роста означает именно численность, соответствующую «емкости угодий»), -стратегия означает «отбор на качество» — повышение адаптивности и устойчивости, а г-стратегия — «отбор на количество» через компенсацию неизбежно больших потерь высоким репродуктивным потенциалом; это —поддержание устойчивости популяции через быструю смену составляющих ее особей. Этот тип стратегии более свойствен мелким животным с большой нормой гибели и высокой плодовитостью (г—коэффициент, отражающий скорость роста популяции). Вицы с г-стратегией легко осваивают местообитания с нестабильными условиями и отличаются высоким уровнем энергозатрат на репродукцию. Выживание этих форм в условиях неблагоприятных абиотических воздействий и сильного пресса конкуренции определяется высоким репродуктивным потенциалом, позволяющим быстро восстановить любые потери в популяции.[ ...]
Рисунки к данной главе:
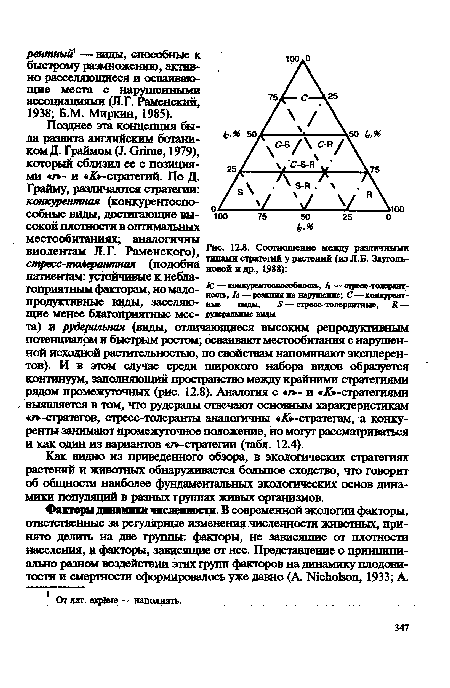
| Соотношение между различными типами стратегий у растений (из JI.E. Зауголь-новой и др., 1988) |
 |
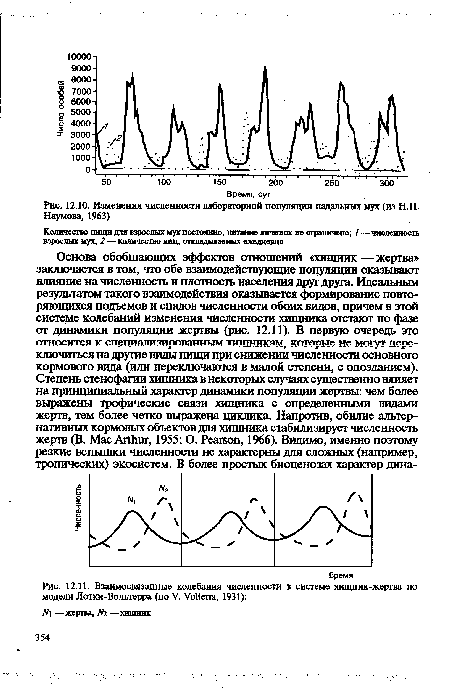
| Изменения численности лабораторной популяции падальных мух (из Н.П. |
 |
| Взаимосвязанные колебания численности в системе хшцник-жертва по модели Лотки-Вольтерра (по V. УоНепа, 1931) |
|
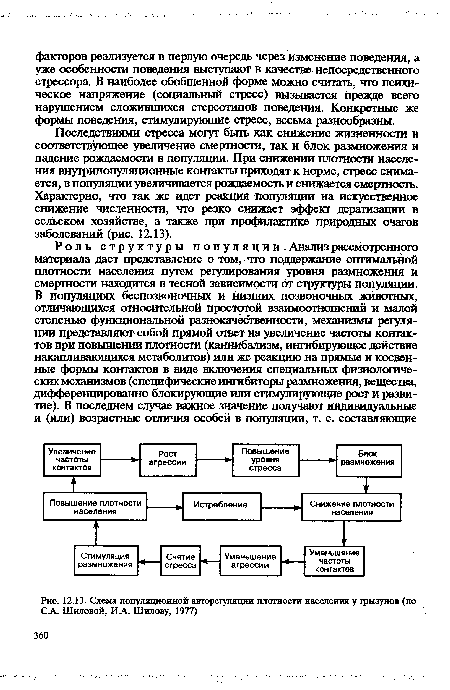
| Схема популяционной авторегуляции плотности населения у грызунов (по СА. Шиловой, ИА. Шилову, 1977) |
 |