Численное соотношение различных категорий организмов в составе населения рассматривается как демографическая структура популяции. При этом в первую очередь имеется в виду соотношение половых и возрастных групп; изменения этих показателей существенным образом влияют на темпы репродукции, а соответственно на общую численность популяции и ее изменения во времени.[ ...]
Возрастные аспекты ценопопуляций растений. В составе ценопопуляций возрастая структура выражена несколькими периодами, включающими ряд определенных возрастных состояний организмов (табл. 12.1).[ ...]
В реальных популяциях границы календарного возраста двух смежных возрастных состояний в какой-то степени перекрываются. Это определяется тем, что индивидуальный ход онтогенеза у отдельных организмов не совпадает и конкретные особи могут достигать определенного возрастного состояния в разные календарные сроки. Функционально же особи данного возрастного состояния сходны. Набор возрастных состояний в популяции определяет интенсивность репродукции, захвата пространства, процессов самоизреживания и т. п. В целом в зависимости от возрастного состава (возрастного спектра) популяция характеризуется определенными свойствами, отражающими ее онтогенетическое состояние как целостной системы.[ ...]
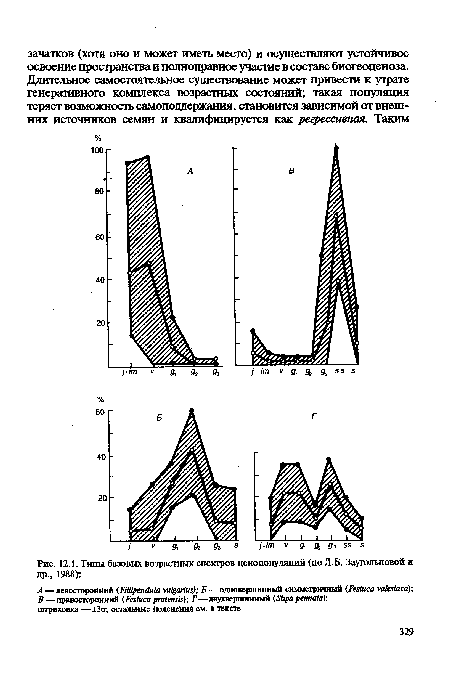
В нормальных дефинитивных (достигших равновесного состояния) ценопопуляциях характер базового возрастного спектра определяется особенностями биологии вида (в частности, общей продолжительностью онтогенеза и длительностью отдельных возрастных состояний), способами самоподцержания, способностью формировать почвенный запас семян и др. Все эти особенности накладывают свой отпечаток на соотношение различных возрастных групп в составе популяции разных видов (рис. 12.1).[ ...]
Возрастная структура популяций животных. Онтогенетические отличия морфологии, физиологии и функциональной роли в популяции свойственны и возрастным группам в популяциях животных. Особенно отчетливо возрастные отличия проявляются у видов, развитие которых проходит с метаморфозом, включающим одну или несколько предимагинальных фаз. В этом случае отдельные возрастные группы могут коренным образом отличаться по особенностям биологии и занимать разное положение в структуре биоценоза. Не представляет исключения и такой вариант развитая, при котором разные стадии онтогенеза обитают в различных средах (например, стрекозы или амфибии).[ ...]
У видов, размножающихся раз в году, когорты выражены четко, а общий спектр возрастного состава зависит от сроков достижения половозрелости и от общей продолжительности жизни, свойственной данному виду. Так, у майских хрущей МеЫоШМ размножение происходит один раз за сезон, после чего самки погибают. При таком ходе онтогенеза понятия «когорта» и «генерация» совпадают и популяция состоит из четырех генераций соответственно четырехлетнему сроку развития обитающих в почве личинок. В других случаях когорты (особи одного времени рождения) могут состоять из представителей одной генерации (последовательные выводки одной когорты родителей), а могут иметь более сложный состав. Так, у мелких грызунов во второй половине репродуктивного сезона в размножение вступают зверьки рождения данного года; соответственно «осенние» когорты состоят из повторных выводков перезимовавших животных и из потомства их детей, т. е. включают по меньшей мере представителей двух поколений (генераций).[ ...]
Как и в популяциях растений, интенсивность размножения и темпы роста популяции в каждый данный момент опред еляются долей особей, находящихся в возрасте активной репродукции; процент неполовозрелых животных в составе популяции отражает потенциальные возможности воспроизводственной функции на ближайшее будущее. Таким образом, знание возрастного состава популяции на фоне видоспецифических сроков развития и созревания может быть основой прогнозирования темпов роста популяций экономически важных видов.[ ...]
Для очень многих видов характерна повышенная смертность в младших возрастных группах (или в предимагинальных стадиях развития). У таких видов кривая выживания демонстрирует резкое падение в облает младших возрастов, которое вскоре сменяется постепенным понижением, отражающим низкую и относительно равномерную смертность животных, переживших «критический» возраст (рис. 12.3, III). При равномерном распределении смертности по возрастам, т. е. в случае независимости причин смертности от специфических возрастных свойств, характер выживания в идеале представляется в виде диагонально снижающейся прямой линии (рис. 12.3, II); приближающийся к этому типу характер выживания свойствен в первую очередь видам, развитие которых идет без метаморфоза при достаточной степени самостоятельности и устойчивости рождающегося потомства, хотя идеально равномерной смертности, по-видимому, не существует2. Во многих случаях видовая кривая выживания характеризуется комбинацией разных частей теоретических кривых (AM. Гиляров, 1990).[ ...]
Анализ параметров выживания и смертности в разных возрастных группах открывает возможность расчета ожидаемой продолжительности жизни особей данной возрастной когорты. Составленные по основным демографическим параметрам таблицы выживания (life tables) могут служить основой анализа и прогнозирования популяционной динамики (табл. 12.2). При этом анализ возрастной динамики может быть основан на последовательных учетах численности отдельных возрастных когорт (как в приведенной таблице) или на статистическом анализе всех возрастных групп, существующих в данной популяции в период наблюдений (AM. Гиляров, 1990).[ ...]
Впрочем, в несбалансированных социальных условиях резко возрастает детская смертность и кривая выживания принимает иной вид.[ ...]
Рисунки к данной главе:
| Типы базовых возрастных спектров ценопопуляций (по Л.Б. Заугольновой и др., 1988) |
 |
| Динамика возрастной структуры популяции рыжих полевок в Ильменском заповеднике (по Г.В. Оленеву, 1983) |
 |
| Схема генетического механизма определения пала на примере слияния гамет у млекопитающих |
 |