Генетическая структура популяции определяется прежде всего богатством популяционного генофонда, включающего как общие видовые свойства, так и особенности, возникшие в порядке приспособления популяции к конкретным условиям ее существования. Этот аспект генетической структуры включает и степень индивидуальной изменчивости (генетический полиморфизм) по комплексу признаков.[ ...]
С другой стороны, важной особенностью генетической структуры оказывается сложность генома каждой отдельной особи, характеризующаяся степенью гетерозиготности по множеству свойств. С. С. Четвериков еще в 1926 г. показал возможность длительного сохранения мутаций (даже инадаптивных) в гетерозиготном состоянии. Он образно говорил о том, что «вид, как губка, впитывает в себя гетерозиготные геновариации, сам оставаясь при этом все время внешне (фенотипически) однородным» (С.С. Четвериков, 1965, с. 48).[ ...]
Эволюционный и экологический аспекты изменчивости. Популяция — элементарная единица эволюционного процесса; в этом аспекте большой интерес представляет собой видовая специфика свойств организма, преобразование генофонда популяции под давлением отбора и некоторых специфических генетических механизмов, часть из которых непосредственным образом связана с особенностями экологии популяций (степень их самостоятельности, наличие и характер волн численности, специфика расселения и др.).[ ...]
Однако специфика и степень сложности генофонда определяют не только микроэволюцион-ные процессы, но и успешное существование популяции в разнообразных и динамичных условиях среды. Широкий диапазон индивидуальной изменчивости лежит в основе устойчивости популяций при отклонениях условий от их средних, типичных характеристик. Чем генетически более разнородна популяция, чем менее специализированы геноа-даптации, тем выше экологическая пластичность популяции, что выгодно как в микроэволю-ционном плане, так и при повседневном приспособлении к текущим условиям среды (О. ваше, 1947; С.С. Шварц, 1972).[ ...]
У животных свободная панмиксия возможна лишь в пределах низших таксонов, не обладающих сложной структурой взаимоотношений, да и в этом случае масштабы свободного обмена генетическим материалом всегда в какой-то мере ограничены неравномерностью условий среды и агрегированностью пространственного распределения особей. В популяциях высших животных панмиксия всегда ограничена вследствие иерархических взаимоотношений и соответствующей неравнозначности особей в проявлении различных форм активности, в том числе участия в размножении.[ ...]
То же свойственно некоторым воробьинообразным. Так, наблюдения за манакинами Chiroxiphia linearis в тропических предгорных лесах Коста-Рики показали, что из 85 самцов на токовищах в копуляциях участвовали всего 8, причем на долю четырех из них пришлось 90 % всех спариваний, а на долю одного — 67 % (D. Me Donald, 1989).[ ...]
Аналогичная картина наблюдается и у некоторых видов млекопитающих. Показано, например, что на лежбищах морских слонов Miro-unga angustirostris секачи-доминанты составляют лишь 4% от всех самцов, но принимают участие в 85 % спариваний и оплодотворяют до 85 % самок; такая же система спариваний характерна для многих других ластоногих.[ ...]
У животных с менее жесткой системой доминирования в размножении участвует соответственно большая доля самцов в популяции. Но элементы «скрытого» сексуального доминирования имеют место и в этом случае. Так, у территориальных животных из размножения исключаются особи, не сумевшие захватить и удержать участок обитания. В некоторых случаях эта часть популяции достаточно велика: у некоторых уток —до 60—80 % всех особей, у австралийской сороки Gymnorhina tibicen — около 75 %, у марабу Leptoptylus crumeniferus —до 80—90 %.[ ...]
Среди обладающих территорией самцов участие в размножении также неодинаково, что в первую очередь связано с их положением в этологической структуре внутрипопуляционных групп. Эта закономерность подтверждена экспериментально: генетический анализ потомства у лабораторных мышей и у гуппи показал достоверное преобладание у молодых животных признаков самцов-доминантов. Полевые наблюдения за длинноклювыми крапивниками Telmatodytespalustris показали, что самки предпочитают образовывать пары с самцами, занявшими лучшие участки и построившими больше гнезд; в результате часть территориальных самцов остается без пары. Аналогичное явление описано для черной каменки Oenante picata-, самцы, занявшие худшие участки, могут остаться холостыми даже при наличии свободных самок. Поскольку качество занятой территории отражает место данного самца в системе взаимоотношений, связь генетического вклада с местом в иерархии прослеживается и в этих случаях.[ ...]
Рисунки к данной главе:
| Варьирование адаптивного признака при изменении величины воздействующего фактора (по И А. Шилову, 1985) |
 |
| Соотношение генотипов в £ « |
 |
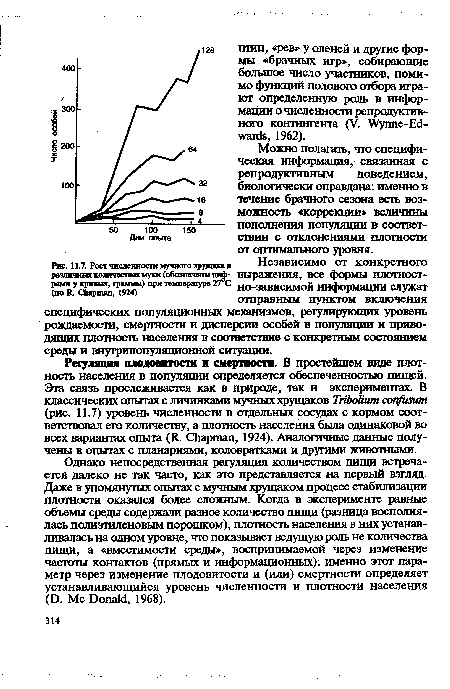
| Рост численности мучного хрущака в Независимо от конкретного различных количествах муки (обозначены Ш Ф- выражения, все формы ПЛОТНОСТ- |
 |
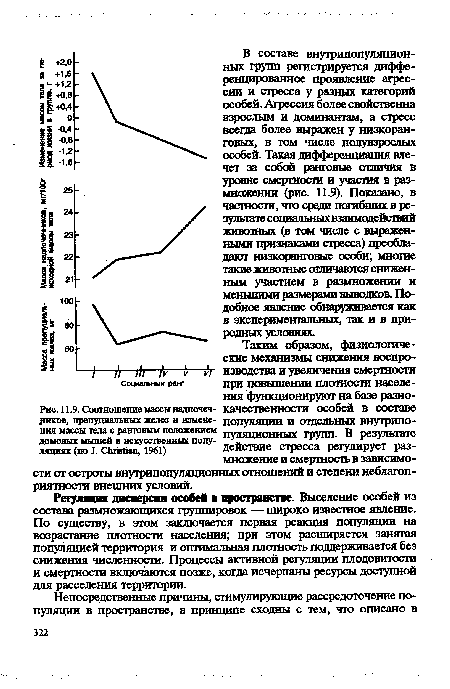
| Соотношение массы надпочечников, препуциальных желез и измене- |
 |