Как было показано выше, конкретные формы пространственного размещения особей в популяциях даже одного и того же вида могут заметно изменяться в зависимости от динамики количества и распределения пищевых объектов (элементов питания), сезонных и непериодических изменений микроклимата, условий, определяющих дальность действия информационных связей, и т. д. Вызываемые этими факторами различия в дистанции между особями и их группировками, размерах последних, характере перемещения в пространстве представляют собой адаптивные варианты видового типа пространственной структуры популяций. Последний же целиком определяется наиболее общими особенностями биологии данного вида.[ ...]
Механизмы «индивидуализации» территории. Биологические преимущества обитания на постоянном участке могут быть реализованы только при условии «индивидуализации» территории, использовании ее лишь постоянно обитающими на ней особями или группами. Как уже было отмечено, особи-резиденты привязаны к территории участка системой знакомых ориентиров, выход за пределы которой вызывает ориентировочную реакцию, служащую стимулом возвращения на участок. Но такой стереотип поведения не гарантирует резидентов от проникновения на их территорию других особей того же вида. Такая гарантия определяется комплексом территориального поведения, свойственного оседлым видам с интенсивным использованием ресурсов.[ ...]
Территориальная агрессия. Наиболее прямая форма территориальных взаимоотношений представлена генетически детерминированным стереотипом агрессивного поведения, направленного на особей своего вида, проникших на территорию участка. Территориальная агрессия в той или иной форме свойственна всем видам, имеющим участки обитания, в том числе и беспозвоночным (некоторые насекомые, ракообразные и др.). Биологически важно, что при территориальных конфликтах победителем (по крайней мере, у позвоночных) в большинстве случаев оказывается особь, на участке которой произошла встреча (особь-резидент, «хозяин» данного участка). Наблюдения, подтверждающие эту закономерность, сделаны для многих видов в естественных популяциях.[ ...]
Весьма показательны опыты, проведенные в естественных условиях с мухоловками-пеструшками Ficedula hypoleuca. Дуплянку с гнездом этих птиц постепенно перемещали, пока она не оказывалась на территории гнездового участка соседней пары. Хозяева участка немедленно начинали атаковать пришельцев и оказывались победителями в завязывающихся схватках. После восстановления исходного положения передвигали гнездовье победителей на территорию ранее побежденной пары. Как только граница их гнездового участка была нарушена, вновь начинались схватки, но при этом активной стороной в инициации агрессии выступали птицы, территория которых подверглась вторжению. Они же и одерживали победу. Если обе дуплянки сближались на «нейтральной» (находящейся вне пределов обоих участков) территории, агрессивных взаимодействий между птицами не возникало (К.Н. Благосклонов, 1962, 1964).[ ...]
Биологический смысл детерминированности победы резидентов в территориальных схватках ясен — в результате этого явления происходит стабилизация системы участков со всеми вытекающими отсюда преимуществами для каждой оседлой особи и для популяции в целом. Механизмы такого пути индивидуализации территории изучены слабо. Можно полагать, что в основе предопределенности решения территориальных конфликтов лежат различия в стереотипах поведения у особей-резидентов и у животных, оказавшихся на незнакомой территории. У первых доминирует врожденный стереотип территориальной агрессии, подкрепленный всей, системой знакомых ориентиров, среди которых появляется объект нападения. У «чужаков» на незнакомой территории развивается ориентировочная реакция — «специфическая целостная деятельность организма, затормаживающая все остальные деятельности» (П.К. Анохин, 1958), которая ингибирует агрессию, и при нападении противника легко переходит в пассивно-оборонитель-ную реакцию. Различная «ориентация» высшей нервной деятельности и решает исход конфликта в пользу «хозяина» участка.[ ...]
Эта гипотеза подтверждается рядом полевых и лабораторных наблюдений. Уже упомянутые исследования А.Н. Солдатовой показывают, что у сусликов, попавших на незнакомую территорию, резко меняется поведение: они передвигаются неуверенно, часто поднимаются «столбиком», демонстрируя все признаки ориентировочного рефлекса. Аналогичным образом ведут себя на незнакомой территории многие другие грызуны. Если создается обстановка, при которой ориентировочный рефлекс возникает и у резидента, шансы партнеров выравниваются. В такой ситуации детерминированность победы определяется иными факторами, а иногда снижается и общий уровень агрессии. Так, в опытах с трехиглой колюшкой Gasterosteus aculeatus было установлено, что агрессивность самца снижается, если увеличивается число одновременно присутствующих противников (J. Van den Assem, 1970). Аналогичные данные получены в опытах с землеройками, грызунами и некоторыми другими животными. Можно полагать, что одновременное присутствие нескольких соперников рассредоточивает внимание резидента, вызывает у него ориентировочную реакцию и этим как бы уравновешивает положение всех особей.[ ...]
По всей вероятности, этот механизм не единственный. У пойки-лотермных животных, например, важную роль в исходе схватки играет температура тела, определяющая активность животного. Показано, что в схватках стенных ящериц Podareis muralis в норме побеждают более крупные самцы, но это правило нарушается, если менее крупный соперник имеет более высокую температуру тела. Однако и в этом случае статус резидента увеличивает шансы на победу (L. Edsman, 1989).[ ...]
Все эти и многие другие данные подтверждают, что победа «хозяев» участков — закономерное явление, биологический смысл которого заключается в стабилизации сложившейся пространственной структуры популяции. Агрессивное поведение эффективно способствует стабилизации системы участков обитания. В полевых опытах с краснокрылыми трупиалами Agelaius phoeniceus отмечено, что если территориальный самец не проявлял агрессии (не нападал на предъявленное чучело трупиала) или агрессия его была выражена слабо, то соседние самцы достоверно чаще расширяли свою территорию за его счет, проявляя таким образом способность оценивать возможность агрессии со стороны соседа по его поведению (S. Freeman, 1987).[ ...]
Рисунки к данной главе:
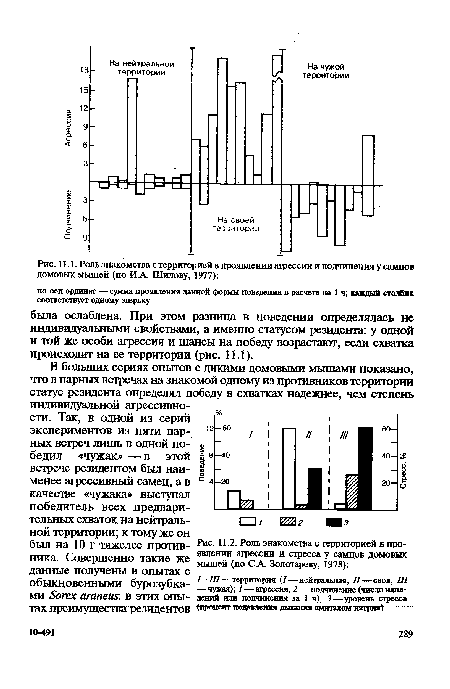
| Роль знакомства с территорией в проявлении агрессии и подчинения у самцов домовых мышей (по И А. Шилову, 1977) |
 |
| Роль знакомства с территорией в проявлении агрессии и стресса у самцов домовых мышей (по С А. Золотареву, 1978) |
|
| Изменение уровня стресс-реакции при формировании иерархии в экспериментальных группах самцов белых мышей (по И.Н. Ивашкиной, 1974) |
 |