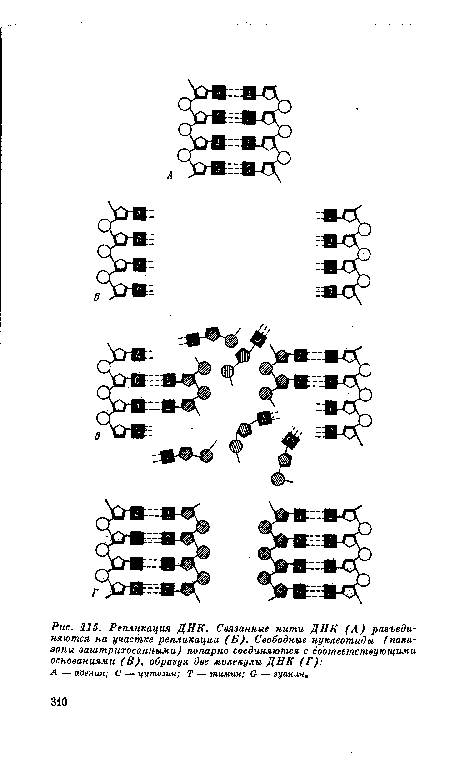
По-видимому, репликация ДНК обеспечивается механизмом, совершенно аналогичным матричной системе наборной машины. Однако матричная модель основывается на химических связях, а не на физическом оттиске. Как показывают химические анализы ДНК, относительное количество азотистых оснований может изменяться, но количество аденина всегда равно количеству тимина, а гуанина — цитозину. Это дает основание полагать, что аденин и тимин и гуанин и цитозин всегда присутствуют парами. Достоверность этого становится очевидной при рассмотрении структуры ДНК. Молекула ДНК обычно состоит из двух цепей, соединенных связями между азотистыми основаниями, при этом аденин всегда спарен и противоположен тимину, а гуанин спарен с цитозином и противоположен ему. Таким образом, если дана последовательность одной цепи, такая же последовательность фиксирована в противоположной цепи. Следовательно, каждая цепь является «матрицей» для другой.[ ...]
Теория репликации ДНК логически вытекает из ее структуры. Разъединение цепей ДНК, очевидно, сопровождается перестройкой добавочных оснований в каждой цепи (рис. 115).[ ...]
А — аденин; С — цитозин; Т — тимин; G — гуанин.[ ...]
А — авенип; С — цитовин; Т — тимип; О — гуанин; и — урацил.[ ...]
Основное различие между бесполым и половым размножением — это разница между двумя типами деления клеток — митозом и мейозом, в характере распределения хромосом. Хромосомы размещаются в ядре, имеют линейную структуру и содержат гены, переносчики генетической информации.[ ...]
В митозе деление хромосом и клетки происходит одновременно. Хромосомы делятся продольно, каждая половинка движется к противоположным концам клетки. Образовавшиеся в результате такого деления две дочерние клетки получают совершенно одинаковое число и тип хромосом. Остальные элементы строения клетки также равномерно делятся между дочерними клетками.[ ...]
В мейозе (последовательные деления клеток при формировании гамет) число хромосом уменьшается вдвое. Хромосомы в обычной вегетативной клетке высших растений, как правило, располагаются парами, образуя соматическое число. Обе хромосомы в паре (гомологи) морфологически подобны и содержат однотипные гены, хотя в результате некоторых отклонений (мутаций) гены внутри пары могут оказаться неидентичными. Генетическая изменчивость живых организмов обусловливается различными сочетаниями генов. Половой процесс — э о механизм, обеспечивающий такую перегруппировку и рекомбинацию генетических факторов, которая позволяет организмам приспосабливаться к изменяющимся условиям среды. Перегруппировка генетических факторов между и внутри хромосом совершается путем мейоза; рекомбинация завершается оплодотворением (см. гл. 10).[ ...]
По существу мейоз представляет собой два деления, в которых клетки делятся дважды, а хромосомы только один раз. Это приводит к образованию четырех клеток, каждая из которых имеет гаплоидное число хромосом, т. е. половинный набор хромосом соматических клеток. Каждая из этих четырех клеток потенциально является гаметой. Оплодотворение (слияние двух гамет) восстанавливает диплоидное число хромосом.[ ...]
Основная разница между митозом и мейозом наблюдается при первом делении. При мейозе, как и при митозе, хромосомы явно удваиваются, но это последнее деление хромосом, хотя клетки будут делиться еще раз. В отличие от митоза две гомологичные хромосомы, теперь явно удвоенные, спариваются (синапсис) по всей своей длине. Притяжение между удвоенными хромосомами сменяется отталкиванием, и каждая удвоенная хромосома пары передвигается к противоположному полюсу. В результате на каждом конце клетки сосредоточивается половина от «целого» числа хромосом.[ ...]
Очевидно, при слиянии хромосом при первом делении происходит фактический обмен сегментов (кроссоверы) между гомологичными хромосомами (обмен включает в себя хроматиды — половинки хромосом). Точный механизм обмена хромосомного материала до сих пор полностью не раскрыт. Следовательно, общим результатом первого деления является не только уменьшение числа хромосом, но и перестройка сегментов между гомологичными хромосомами. Второе деление хромосом, следующее сразу за первым, протекает подобно митотическому делению, но два обособившихся дочерних ядра из-за кроссоверов не могут быть тождественными.[ ...]
Рисунки к данной главе:
Вернуться к оглавлению