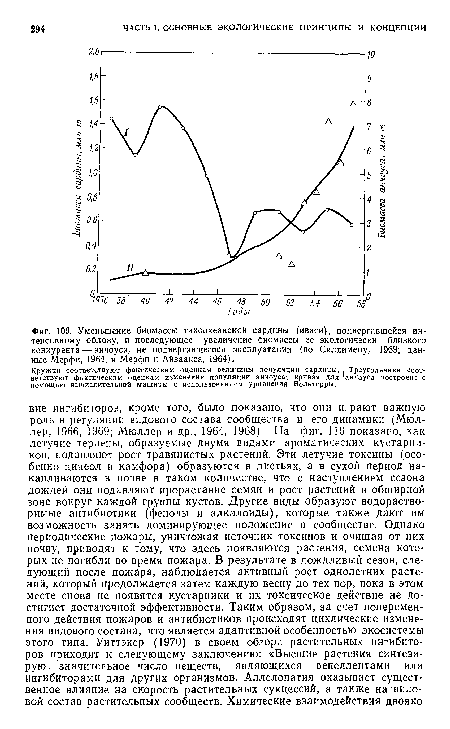
Основная особенность отрицательного взаимодействия популяций заключается в том, что при их синхронной эволюции в стабильной экосистеме степень отрицательного влияния уменьшается. Иными словами, естественный отбор стремится уменьшить отрицательные влияния или вообще устранить взаимодействие популяций, поскольку продолжительное и сильное подавление популяции добычи или хозяина популяцией хищника или паразита может привести к уничтожению одной из них или обеих. Жесткая конкуренция чаще всего отмечается там, где контакт между популяциями установился недавно (в начале объединения двух популяций или когда в экосистеме произошли значительные или внезапные, возможно временные, изменения, например под влиянием деятельности человека). Именно поэтому, в частности, внезапное или непродуманное вмешательство человека в природу нередко приводит к эпидемическим вспышкам.[ ...]
Объективный подход к вопросу о паразитизме и хищничестве оказывается нелегким делом. Все мы испытываем естественное отвращение к паразитам, будь то бактерии или ленточные глисты. Люди настроены и против всех хищников, хотя человек — самый типичный хищник (и основной виновник природных эпидемий). К тому же обвинительный приговор хищнику часто выносится наобум, без полной уверенности в том, что он действительно вредит интересам человека. «Волк хорош только мертвым» — эта широко распространенная точка зрения, как мы увидим далее, оказывается весьма далекой от истины.[ ...]
Переходим к наиболее важному обобщению, а именно что отрицательные взаимодействия со временем становятся менее заметными, если экосистема достаточно стабильна и ее пространственная структура обеспечивает возможность взаимного приспособления популяций. Как отмечалось в разд. 9, в простых популяциях типа паразит — хозяин и хищник — жертва, введенных в экспериментальные микроэкосистемы, наблюдаются сильные колебания численности, характеризующиеся определенной вероятностью затухания. В модельных системах типа хищник— жертва, описываемых уравнением Лотки—Вольтерры, если в уравнение не введены дополнительные члены, характеризующие действие факторов самоограничения численности, то колебания происходят непрерывно и не затухают (см. Левонтин, 1969). Пиментел (1968; см. также Пиментел и Стоун, 1968) экспериментально показал, что такие дополнительные члены могут отражать взаимные адаптации или генетическую обратную связь. Как видно на фиг. 105, сильные колебания численности происходят в том случае, когда хозяина (комнатную муху) и паразита (осу) впервые помещают вместе в ограниченную культуру. Когда же новые культуры создавали из особей, ранее на протяжении двух лет совместно существовавших в культуре, где их численность подвергалась значительным колебаниям, оказалось, что у них выработался экологический гомеостаз, при котором каждая из популяций была «подавлена» другой в такой степени, что оказалось возможным их сосуществование при более стабильном равновесии.[ ...]
Однако в реальном мире обстоятельства могут складываться таким образом, что взаимная адаптация в новых ассоциациях не возникает. Поэтому всегда существует опасность, что отрицательная реакция может стать необратимой и хозяин будет элиминирован. История с грибным заболеванием каштана в Америке служит примером такого взаимодействия популяций, которое с равной вероятностью может закончиться и адаптацией, и вымиранием, и человеку ничего не остается, как быть лишь пассивным наблюдателем происходящего.[ ...]
В самом деле, если составить список болезней, паразитов и насеко-■мых-вредителей, причиняющих наибольший убыток сельскому хозяйству или наиболее патогенных для человека, то окажется, что многие из них недавно занесены в новые для них местности и нашли нового хозяина или добычу.[ ...]
Урок из всего этого таков: необходимо избегать создания новых отрицательных взаимодействий, а если они возникли, стараться по возможности сдерживать их.[ ...]
Хотя с экологической точки зрения хищничество и паразитизм очень сходны, между их крайними формами — крупным хищником и мелким внутренним паразитом — имеются весьма существенные различия, касающиеся не только размеров. Паразитические и патогенные организмы обычно обладают более высоким биотическим потенциалом, чем хищники. Их строение, обмен, отношение к хозяину и жизненный цикл часто более специализированы, что связано со специфичностью условий их среды и с переселением от одного хозяина к другому.[ ...]
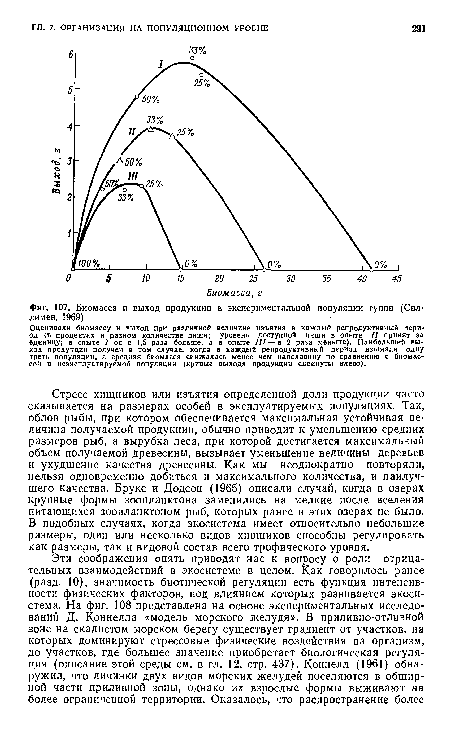
К анализу проблемы можно подойти и экспериментально, создавая опытные популяции в микроэкосистемах. Одна из таких экспериментальных моделей представлена на фиг. 107. Аквариумную рыбку гуппи (1еЫз1ез гейси1аЫз) использовали для имитации популяции промысловой рыбы, облавливаемой человеком. Можно видеть, что максимальный устойчивый выход продукции получали в том случае, когда в каждый репродуктивный период изымалась одна треть популяции, что приводило к уменьшению равновесной плотности до величины, составлявшей несколько меньше половины плотности необлавливавшейся популяции. Эксперимент показал также, что эти отношения независимы от предельной емкости системы, которая поддерживалась на трех разных уровнях путем изменения количества пищи.[ ...]
Модели, состоящие из одного вида, часто слишком упрощают реальные отношения, поскольку в них не учитывается роль конкурирующих видов; между тем плотность последних может увеличиваться из-за уменьшения плотности изымаемого вида, и, кроме того, они используют пищу или другие ресурсы, необходимые для существования изучаемого вида. «Верховному» хищнику, например человеку (или наиболее эффективному пастбищному животному, например корове), очень легко повлиять на конкурентное равновесие, что приведет к замещению эксплуатируемого вида на другой, непригодный для данного хищника или данного пастбищного животного. Случаи таких сдвигов (как будет отмечено ниже) все множатся по мере того, как активизируется рыболовная и охотничья и т. п. деятельность человека. Это дает основание высказать следующее утверждение, которое следует рассматривать и как призыв, и как предупреждение: эксплуатируемые для нужд человека системы, представленные одним видом, равно как и системы монокультур (например, сельскохозяйственные монокультуры), неустойчивы по своей природе, ибо в условиях стресса они сильно уязвимы для конкурентов, возбудителей болезней, паразитов, хищников и других факторов, оказывающих отрицательное влияние. Как отмечалось в гл. 3, оптимальная продуктивность может быть меньше максимальной, если принять во внимание затраты, необходимые для поддержания «порядка» в нестабильных системах.[ ...]
Оценивали биомассу и выход при различной величине изъятия в каждый репродуктивный пери« од (в процентах н разном количестве пнщн; уровень доступной пищи в опыте II принят за единицу; в опыте / он в 1,5 раза больше, а в опыте III — в 2 раза меньше). Наибольший вы ход продукции получен в том случае, когда в каждый репродуктивный период изымали одну треть популяции, а средняя биомасса снижалась менее чем наполовину по сравнению с биомассой в неэксплуатнруемой популяции (кривые выхода продукции сдвинуты влево).[ ...]
Рисунки к данной главе:
| Американские каштаны, пораженные паразитическим грибом (Аппалачские горы, Джорджия) |
 |
| Биомасса и выход продукции в экспериментальной популяции гуппи (Сил-лимен, 1969). |
 |
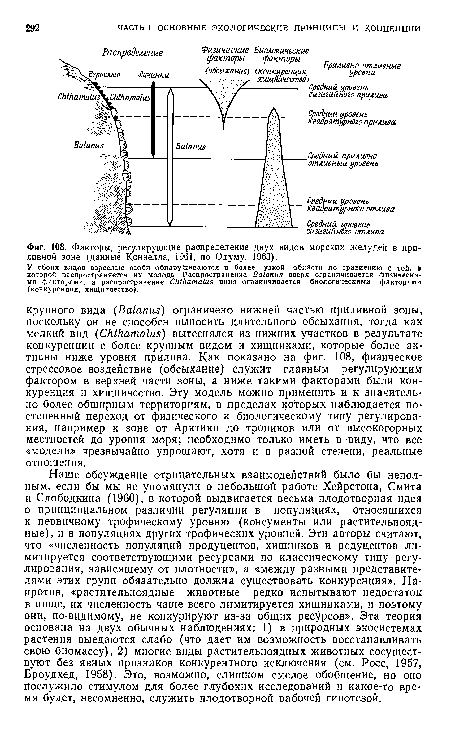
| Факторы, регулирующие распределение двух видов морских желудей в приливной зоне (данные Коннелла, 1961, по Одуму, 1963). |
 |