И еще одно замечание. Обратите внимание на то, что слово «вредный» ни разу не употреблялось при описании отрицательных взаимодействий. Конкуренция и хищничество снижают скорость роста популяции, подвергающейся воздействию, но этот эффект не обязательно вреден, если рассматривать его с точки зрения выживания популяции на протяжении длительного Бремени или с точки зрения ее эволюции. Отрицательные взаимодействия могут ускорять естественный отбор, приводя к возникновению новой адаптации. Как мы уже видели, хищники и паразиты часто полезны для популяций, не имеющих механизмов саморегуляции, для предотвращения перенаселения, следствием которого могло бы быть самоуничтожение (см. разд. 8).[ ...]
Наконец, положение, что в ходе развития популяций наблюдается тенденция к уменьшению отрицательных взаимодействий и увеличению положительных, сформулированное в начале этого раздела, имеет чрезвычайно важное значение для человека. Оно будет обсуждаться в последующих разделах.[ ...]
О межвидовой конкуренции написано очень много и экологами, и генетиками, и эволюционистами. В большинстве случаев слово «конкуренция» применяется для тех ситуаций, в которых отрицательные влияния возникают вследствие уменьшения ресурсов, используемых обоими видами. Тогда непосредственные взаимодействия, такие, как взаимное хищничество или выделение вредных веществ, должны быть отнесены к иной категории явлений (как сделано в табл. 30), хотя для их обозначения и нет общепринятого термина. Межвидовые химические посредники (медиаторы) (или аллелохимические вещества, см. Уиттэкер, 1970), благодаря которым один вид имеет преимущества в конкуренции с другим, названы аллелопатическими веществами (см. стр. 34 и 286) или антагонистическими алломонами (Браун и др., 1970). Я полагаю, что такие слова, как «конкуренция», «сообщество», «популяция», широко используемые не только в науке, но и в обычном языке, следует точно определить (что мы и сделали), но при этом сохранить за ними их широкий смысл. Будет значительно меньше путаницы, если за основным термином оставить широкое значение, а затем, пользуясь соответствующими прилагательными или модифицирующими смысл термина фразами, провести «дробление» для его применения в более узких значениях. Например, мы можем говорить о конкуренции за ресурсы, «антибиотической» конкуренции или конкуренции за свет. Мы могли бы охарактеризовать конкуренцию также количественно, по степени подавляющего влияния, как это сделал Филип (1955), используя метод моделирования. Таким образом, в общем следует избегать специальных терминов, пользуясь ими лишь в тех случаях, когда необходимо охарактеризовать какое-либо очень специальное явление. .[ ...]
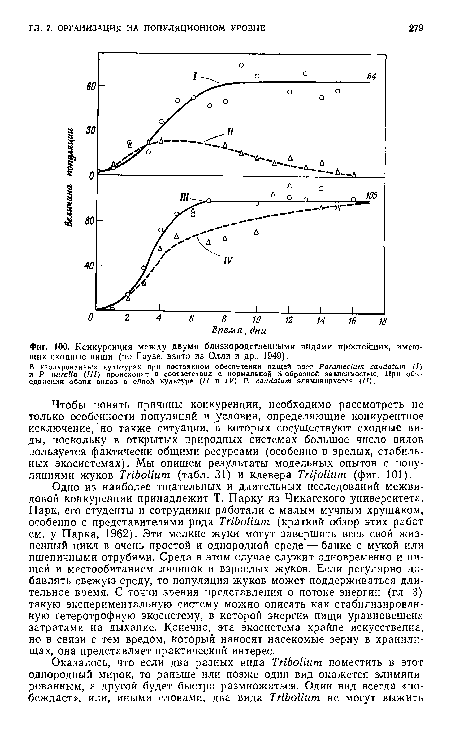
Чтобы понять причины конкуренции, необходимо рассмотреть не только особенности популяций и условия, определяющие конкурентное исключение, но также ситуации, в которых сосуществуют сходные виды, поскольку в открытых природных системах большое число видов пользуется фактически общими ресурсами (особенно в зрелых, стабильных экосистемах). Мы опишем результаты модельных опытов с популяциями жуков Tribolium (табл. 31) и клевера Trifolium (фиг. 101).[ ...]
Одно из наиболее тщательных и длительных исследований межвидовой конкуренции принадлежит Т. Парку из Чикагского университета. Парк, его студенты и сотрудники работали с малым мучным хрущаком, особенно с представителями рода Tribolium (краткий обзор этих работ см. у Парка, 1962). Эти мелкие жуки могут завершить весь свой жизненный цикл в очень простой и однородной среде — банке с мукой или пшеничными отрубями. Среда в этом случае служит одновременно и пищей и местообитанием личинок и взрослых жуков. Если регулярно добавлять свежую среду, то популяция жуков может поддерживаться длительное время. С точки зрения представления о потоке энергии (гл. 3) такую экспериментальную систему можно описать как стабилизированную гетеротрофную экосистему, в которой энергия пищи уравновешена затратами на дыхание. Конечно, эта экосистема крайне искусственна, но в связи с тем вредом, который наносят насекомые зерну в хранилищах, она представляет практический интерес.[ ...]
Но достаточно лабораторных примеров. Совершенно очевидно, что в лабораторных экспериментах степень перенаселенности может быть весьма значительной, что приводит к чрезмерной конкуренции. В полевых условиях межвидовая конкуренция была подробно изучена у растений; результаты этих исследований позволили прийти к выводу (в настоящее время этот вывод является общепринятым), что она служит важным фактором сукцессии видов (гл. 9). Кивер (1955) описала интересный случай, когда залежь первого года была почти целиком занята чистыми травостоями высокого сорняка, в дальнейшем его постепенно вытеснил отсюда другой вид, ранее в этой местности не известный. Эти два вида, хотя и относились к разным родам, имели весьма сходный жизненный цикл (время цветения, время созревания семян) и биологические особенности и поэтому сильно конкурировали между собой.[ ...]
Григгс (1956) провел интересное исследование конкуренции растений на скалистых горных вырубках, где большинство видов растет на отдельных кочках. Кочки одного вида часто оккупируются другим видом. Григгсу удалось сгруппировать виды по их способности занимать другие кочки. Эта «конкурентная лестница», как оказалось, отличалась от порядка сукцессии, поскольку между способностью к вторжению в область существования другого вида и способностью к его полному вытеснению нет полной корреляции.[ ...]
Если преобладает внутривидовая конкуренция, то вид занимает ббльшую территорию, распространяясь и на меиее благоприятные (крайние) участки ареала; если же межвидовая конкуренция значительна, то вид существует на меньшей территории с оптимальными условиями.[ ...]
Рисунки к данной главе:
| Конкуренция между двумя близкородственными видами простейших, имеющих сходные ниши (по Гаузе, взято из Олли и др., 1949). |
 |
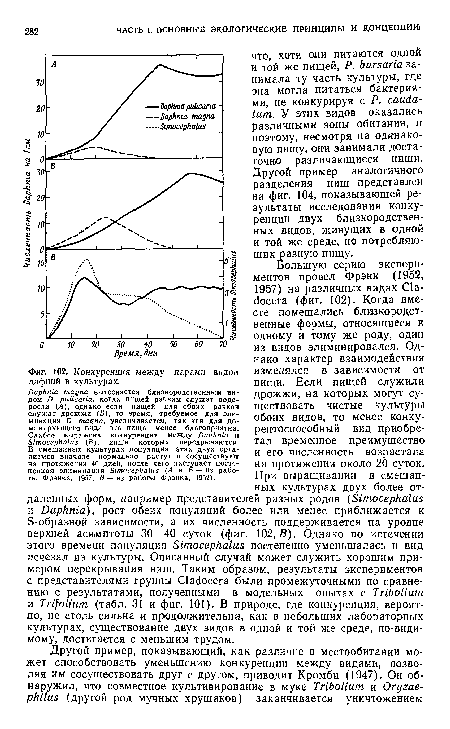
| Конкуренция между парами видов дафний в культурах. |
 |
| Влияние конкуренции на распределение местообитаний у птиц (по Свердсо-ну, 1949, с изменениями). |
 |