Как уже неоднократно подчеркивалось, в своем широком варианте концепция лимитирующих факторов не ограничивается физическими факторами, поскольку биологические взаимоотношения («взаимодействия», или «биологические факторы», среды) не менее важны как регуляторы распределения и численности организмов в природе. Однако биологические факторы удобнее будет рассмотреть в последующих главах, трактующих о популяциях и сообществах; здесь же мы рассмотрим физические и химические аспекты среды. Чтобы изложить все известное по этому вопросу, потребовалась бы целая книга, а это не входит в задачу нашего обзора экологических принципов. Кроме того, рассмотрение подробностей отвлекло бы нас от основной цели — получить общую картину предмета экологии. Поэтому мы лишь кратко перечислим основные моменты, заслуживающие изучения с точки зрения экологов.[ ...]
Степень изменчивости температуры крайне важна для экологии. Температура, колеблющаяся от 10 до 20 °С со средней 15°, не обязательно действует на организмы так же, как постоянная температура 15 °С. Обнаружено, что организмы, которые обычно в природе подвергаются воздействию переменных температур (что имеет место в большинстве умеренных районов), как правило, хуже переносят постоянную температуру. Так, в своем первом исследовании в данной области Шел-форд (1929) обнаружил, что яйца и личинки или куколки яблонной плодожорки в условиях колеблющейся температуры развиваются на 7 или 8% быстрее, чем при постоянной температуре, равной средней температуре в опыте. В другом эксперименте (Паркер, 1930) с помощью переменной температуры удалось ускорить развитие яиц кузнечика в среднем на 38,6%, а нимф — на 12% по сравнению с развитием при близкой постоянной температуре.[ ...]
Не ясно, связан ли этот ускоряющий эффект с самими колебаниями температуры или с усиленным ростом вследствие временного повышения температуры. Во всяком случае, стимулирующий эффект переменных температур, по крайней мере в умеренной зоне, можно считать четко установленным, и это необходимо подчеркнуть, поскольку лабораторные эксперименты проводятся обычно при постоянной температуре.[ ...]
Так как организмы чувствительны к изменениям температуры и так как температуру легко измерять, ее роль как лимитирующего фактора иногда переоценивают. Подобных выводов надо остерегаться; иногда другие, не измеренные факторы могут быть важнее. О распространенной способности растений, животных и особенно сообществ компенсировать температуру или акклимироваться к ней упоминалось выше. Начинающему экологу можно посоветовать следующее: при изучении конкретного организма или проблемы непременно учитывайте температуру, но не останавливайтесь на этом.[ ...]
Для экологии важны качественные признаки света (длина волны или цвет), интенсивность (действующая энергия в калориях или фут-канделах) и продолжительность воздействия (длина дня). Известно, что и животные, и растения реагируют на изменения длины волны света. Цветовое зрение распространено в разных таксономических группах животных. «пятнисто»; оно, по-видимому, хорошо развито у некоторых видов членистоногих, рыб, птиц и млекопитающих, но у других видов тех же групп оно может отсутствовать (например, из млекопитающих цветовое зрение хорошо развито только у приматов). Интенсивность фотосинтеза несколько варьирует с изменением длины волны света. В наземных экосистемах качественные характеристики солнечного света не настолько изменчивы, чтобы это сильно влияло на интенсивность фотосинтеза, но при прохождении света через воду красная и синяя части спектра отфильтровываются и получающийся зеленоватый свет слабо поглощается хлорофиллом. Однако красные водоросли имеют дополнительные пигменты (фикоэритрины), позволяющие им использовать эту энергию и жить на большей глубине, чем смогли бы зеленые водоросли (см. гл. 12, разд. 5).[ ...]
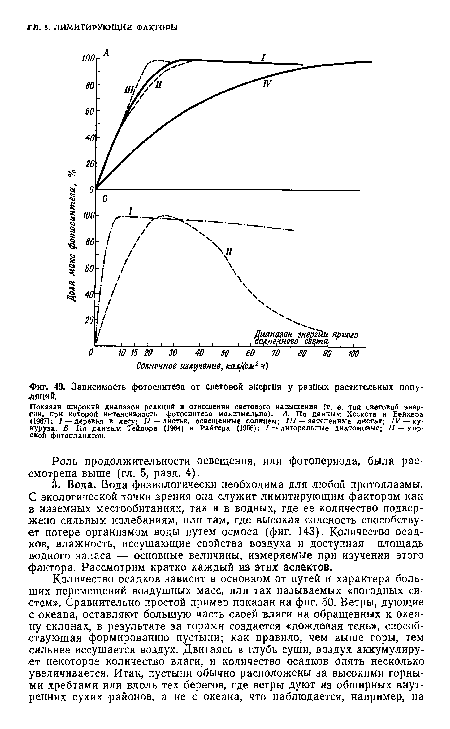
Как показано в гл. 3, интенсивность света (т. е. поступление энергии), падающего на автотрофный ярус, управляет всей экосистемой, влияя на первичную продуктивность. И у наземных и водных растений фотосинтез связан с интенсивностью света линейной зависимостью до оптимального уровня светового насыщения, за которым во многих случаях следует падение фотосинтеза при очень сильных интенсивностях света (Рабинович, 1951; Томас, 1955). На фиг. 49 показаны кривые зависимости фотосинтеза от интенсивности света у нескольких популяций и сообществ. Как и следовало ожидать, компенсация факторов затемняет картину, так как отдельные растения и целые сообщества приспосабливаются к разным интенсивностям света, становясь «адаптированными к тени» (т. е. достигают насыщения при низкой интенсивности) или «адаптированными к солнечному свету» (гл. 3, фиг. 18). Крайние значения светового насыщения показаны на фиг. 49. Диатомовые, живущие в песке пляжей или на илистой литорали, интересны тем, что максимальный фотосинтез достигается у них при интенсивности света менее 5% интенсивности прямого солнечного овета и чистая продукция образуется при интенсивности света менее 1% полной (Тейлор, 1964). Однако при высокой интенсивности света продукция у этих диатомовых подавляется лишь незначительно. Морской фитопланктон также адаптирован к низкой интенсивности света; свет высокой интенсивности очень сильно его подавляет, и в результате максимум продукции в океане приходится обычно не на поверхностный слой, а на слой, лежащий несколько ниже. На другом полюсе — солнцелюбивые хлебные злаки, для которых световое насыщение достигается только при ярком солнечном свете (фиг. 49).[ ...]
Не все согласны с тем, что нормальный яркий солнечный свет также может лимитировать развитие организмов. При высокой интенсивности света фотоокисление ферментов, видимо, ослабляет синтез, а активное дыхание ведет к расходованию продукта фотосинтеза. Особенно ослабляется синтез белка, так что при ярком освещении в продукте повышается процент углеводов. Именно поэтому в тропиках трудно получить высокие урожаи культур, богатых белком.[ ...]
Показан широкий диапазон реакций в отношении светового насыщения (т. е. той световой энергии, при которой интенсивность фотосинтеза максимальна). А. По данным Хескета н Бейкера <1967); I — деревья в лесу; // —листья, освещенные солнцем; /// — затененные листья; IV — кукуруза. Б По данным Тейлора (1964) и Райтера (1956); / — литоральные диатомовые; // — морской фитопланктон.[ ...]
Роль продолжительности освещения, или фотопериода, была рассмотрена выше (гл. 5, разд. 4).[ ...]
В действительности тип биоты определяется не одним количеством осадков, но равновесием между осадками и потенциальной эвапотран-спирацией (потерей воды сообществом через испарение, о чем подробнее рассказывается далее).[ ...]
Рисунки к данной главе:
| Электромагнитный спектр и его видимая область (из Колуэлла и др , 1963) |
 |
| Зависимость фотосинтеза от световой энергии у разных растительных популяций. |
 |


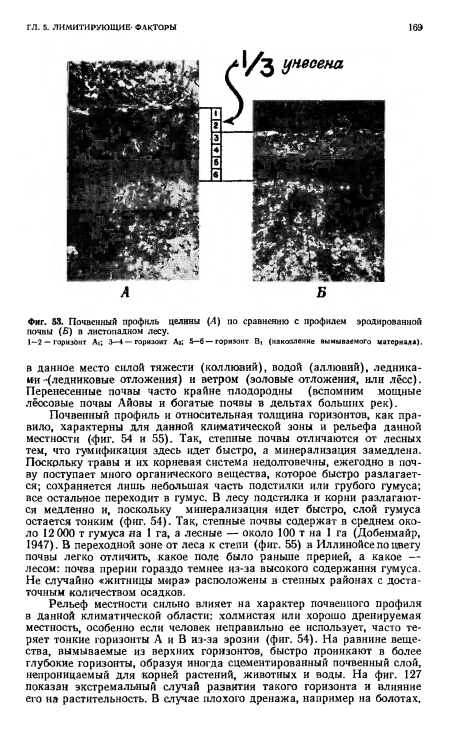
| Почвенный профиль целины (А) по сравнению с профилем эродированной почвы (Б) в листопадном лесу. |
 |
| Упрощенные схемы трех основных типов почвы, характерных для трех биотических зон. |
 |
| Вересковое болото в Англии, выжженное полосами и пятнами, чтобы увеличить продукцию дичи. |
 |
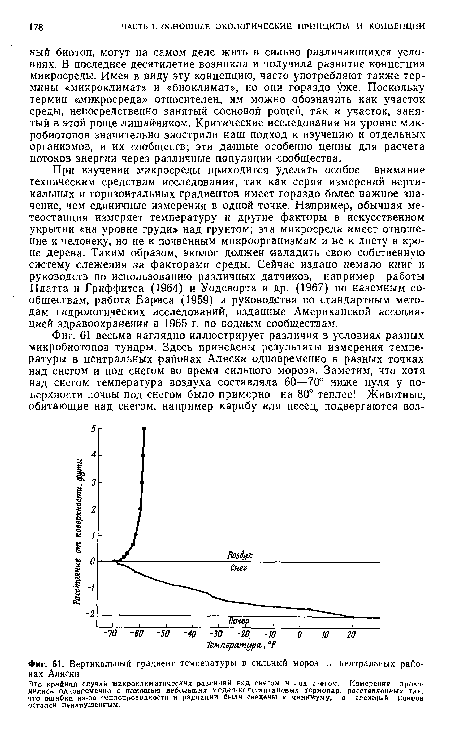
| Вертикальный градиент температуры в сильный мороз в центральных районах Аляски |
 |
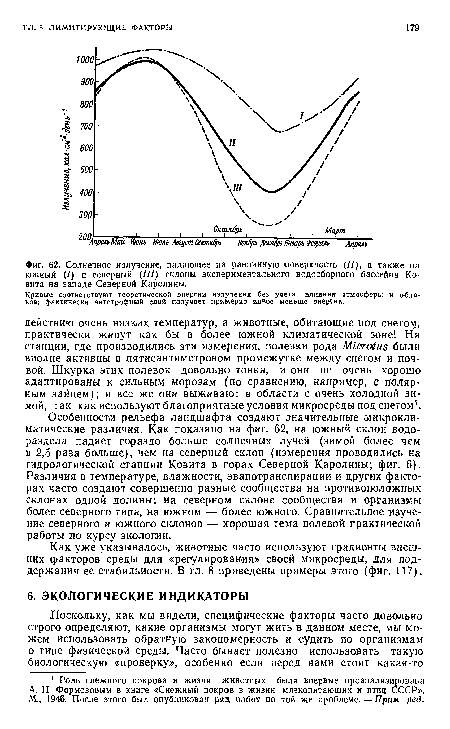
| Солнечное излучение, падающее на равнинную поверхность (//), а также на южный (/) и северный (///) склоны экспериментального водосборного бассейна Ко-внта на западе Северной Каролины. |
 |