Чтобы жить и процветать в тех или иных конкретных условиях, организм должен иметь вещества, необходимые ему для роста и размножения. Основные потребности у разных видов и в разных условиях различны. При «стационарном состоянии» лимитирующим будет то вещество, доступные количества которого наиболее близки к необходимому минимуму. Этот «закон» минимума в меньшей степени приложим к «переходным состояниям», когда быстро измеряются количества, а значит, и эффект многих составляющих.[ ...]
Идея о том, что выносливость организма определяется самым слабым звеном в цепи его экологических потребностей, впервые была высказана в 1840 г. Ю. Либихом, который первым начал изучение влияния разнообразных факторов на рост растений. Он установил, что урожай зерна часто лимитируется не теми питательными веществами, которые требуются в больших количествах, такими, например, как двуокись углерода и вода (поскольку эти вещества обычно присутствуют в изобилии), а теми, которые требуются в малых количествах (например, бор), но которых и в почве мало. Выдвинутый Либихом принцип: «Веществом, находящимся в минимуме, управляется урожай и определяется величина и устойчивость последнего во времени», — получил известность как либиховский «закон» минимума. Многие авторы (например, Тейлор, 1934) расширили это положение, включив в него, помимо питательных веществ, и ряд других факторов, например температуру и время. Чтобы избежать путаницы, лучше, пожалуй, ограничить концепцию минимума, применяя ее, как это делал сам Либих, лишь к химическим веществам (кислороду, фосфору и т. д.), необходимым для роста и размножения организмов; другие же факторы и лимитирующий эффект максимума включить в «закон» толерантности. Обе эти концепции могут быть объединены в общий принцип лимитирующих факторов (см. ниже). Таким образом, «закон» минимума — это лишь один аспект зависимости организмов от среды.[ ...]
Пример двуокиси углерода особенно интересен в свете текущих споров в литературе по поводу загрязнения воды. Споры идут о том, что служит главным лимитирующим фактором и, следовательно, основным веществом, вызывающим процесс «культурной эвтрофикации» в пресных водах, — двуокись углерода или фосфор (Кюнцель, 1969). Поскольку культурная звтрофикация обычно создает в высшей степени неустойчивое состояние с резкими колебаниями (бурные «цветения» водорослей с последующим отмиранием, при котором высвобождаются питательные вещества, вызывающие очередное «цветение»), то нельзя выдвигать на первое место в качестве лимитирующего фактора какое-то одно из веществ. Во время таких колебаний фосфор, азот, двуокись углерода и многие другие вещества могут быстро сменять друг друга как лимитирующие факторы. Следовательно, в таких переходных условиях нет оснований для гипотезы «одного фактора». Чтобы предупредить эвтрофи-кацию, борьба с загрязнениями должна быть направлена не только против органических веществ (дающих СОг и, очевидно, стимулирующие рост другие органические вещества), но и против минеральных веществ, также необходимых для «злокачественного» увеличения продуктивности.[ ...]
Второй вспомогательный принцип касается взаимодействия факторов. Так, высокая концентрация или доступность некоторого вещества или действие другого (не минимального) фактора может изменять потребление минимального питательного вещества. Иногда организм способен заменять, хотя бы частично, дефицитное вещество другим, химически близким Так, в местах, где много стронция, моллюски иногда частично заменяют кальций в своих раковинах стронцием. Показано, что некоторым растениям нужно меньше цинка, если они растут не на ярком солнечном свету, а в тени, в этих условиях имеющееся в почве количество цинка становится уже не лимитирующим.[ ...]
Само присутствие и процветание организма в данном местообитании зависят от целого комплекса условий. Отсутствие или невозможность процветания определяется недостатком (в качественном или количественном смысле) или, наоборот, избытком любого из ряда факторов, уровень которых может оказаться близким к пределам переносимого данным организмом.[ ...]
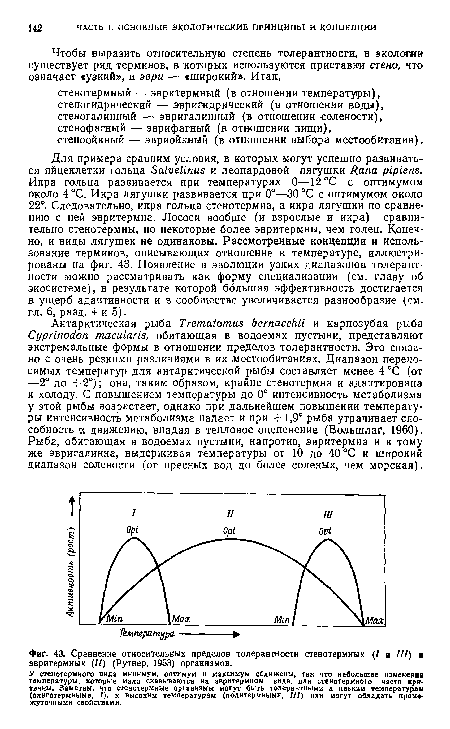
Для примера сравним условия, в которых могут успешно развиваться яйцеклетки гольца Salvelinus и леопардовой лягушки Rana pipiens. Икра гольца развивается при температурах 0—12 °С с оптимумом около 4 °С. Икра лягушки развивается при 0°—30 °С с оптимумом около 22°. Следовательно, икра гольца стенотермна, а икра лягушки по сравнению с ней эвритермна. Лососи вообще (и взрослые и икра) сравнительно стенотермны, но некоторые более эвритермны, чем голец. Конечно, и виды лягушек не одинаковы. Рассмотренные концепции и использование терминов, описывающих отношение к температуре, иллюстрированы на фиг. 43. Появление в эволюции узких диапазонов толерантности можно рассматривать как форму специализации (см. главу об экосистеме), в результате которой большая эффективность достигается в ущерб адаптивности и в сообществе увеличивается разнообразие (см. гл. 6, разд. 4 и 5).[ ...]
У етенотермного вида минимум, оптимум и максимум сближены, так что небольшие изменения температуры, которые мало сказываются на эвритермном виде, для етенотермного часто критичны. Заметим, что стенотермные организмы могут быть толерантными к низким температурам (олнготермными, /), к высоким температурам (полнтермнымн, III) или могут обладать промежуточными свойствами.[ ...]
Конечно, экологическая эффективность не одинакова по всему диапазону; например, усвоение пищи максимально при температуре 20° и солености 15%о (Лоув и Хит, 1969).[ ...]
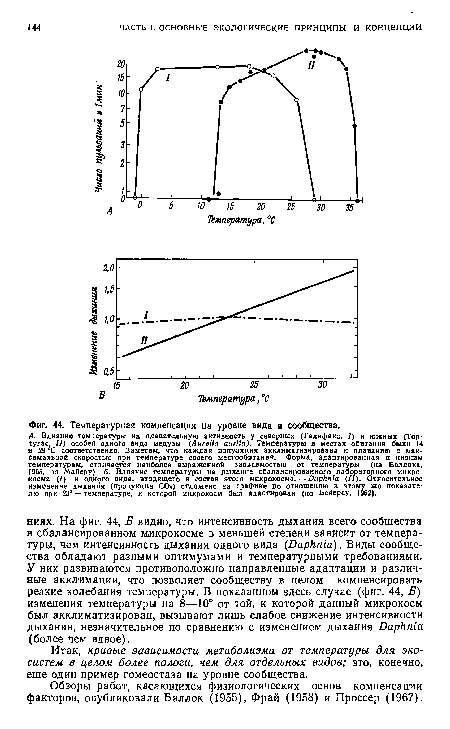
Как уже много раз подчеркивалось, организмы не являются рабами физических условий среды; они приспосабливаются сами и изменяют условия среды так, чтобы ослабить лимитирующее влияние температуры, света, воды и других физических факторов. Такая компенсация факторов особенно эффективна на уровне сообщества, но возможна и на уровне вида. Виды с широким географическим распространением почти всегда образуют адаптированные к местным условиям популяции, называемые экотипами. Их оптимумы и пределы толерантности соответствуют местным условиям. Компенсация в отношении разных участков градиента температуры, света и других факторов может сопровождаться появлением генетических рас (с морфологическими проявлениями или без них) или может быть просто физиологической акклимацией. Удобным методом для проверки того, сопровождается ли появление экотипов генетическим закреплением, служит метод реципрокных пересадок. МакМиллан (1956), например, обнаружил, что злаки, принадлежащие к одному виду и идентичные по всем внешним признакам, при пересадке на экспериментальные участки из разных районов географического ареала по-разному реагировали на свет. В каждом случае они сохраняли приспособленную к исходному району сезонную цикличность (время роста и размножения). В прикладной экологии часто оставляли без внимания возможность генетического закрепления особенностей местных линий, в результате чего интродукция животных и растений часто оканчивалась неудачей, так как вместо приспособленных к местным условиям линий использовались особи из отдаленных областей. Компенсация факторов в местном или сезонном градиенте также может сопровождаться появлением генетических рас, но часто осуществляется за счет физиологической адаптации органов или сдвигов взаимоотношений «фермент—субстрат» на клеточном уровне. Сомеро (1969), например, указывает, что кратковременная температурная компенсация основана на обратной зависимости сродства фермента к субстрату от температуры, а длительная эволюционная адаптация скорее изменяет само это сродство. У животных, особенно у крупных, с хорошо развитой локомоторной способностью, компенсация факторов возможна благодаря адаптивному поведению — они избегают крайностей местного градиента условий. Примеры такой поведенческой регуляции, которая может быть не менее эффективна, чем внутренняя, физиологическая, приведены в гл. 8, разд. 8.[ ...]
Рисунки к данной главе:
| Сравнение относительных пределов толерантности стенотермных (7 и III) и эвритермных (11) (Рутнер, 1953) организмов. |
 |
| Температурная компенсация на уровне вида и сообщества. |
 |