Для определения структуры и функции экосистемы иногда важнее бывает оценить скорости обмена или переноса веществ, нежели количе-ство веществ в данное время в данном месте. Для понимания круговорота веществ и лучшего контроля его человеком необходимо изучить количественно скорость циркуляции и статику явлений. С использованием в экологии разнообразных современных методов, в том числе метода радиоактивных индикаторов, гпектрофотометрии, хроматографии, автоматического управления и дистанционных измерений, появилась возможность измерять скорости циркуляции в довольно крупных экосистемах, например озерах и лесах, и приступить к самой важной задаче — получить количественные сведения о биогеохимических круговоротах в глобальном масштабе.[ ...]
Пруды и озера особенно удобны для исследований, поскольку на протяжении короткого периода времени круговороты биогенных элементов в них могут рассматриваться как независимые. Вслед за первыми опытами Коффина и др. (1949), а также Хатчинсона и Бауэна (1948, 1950) появились многочисленные сообщения о результатах использования радиоактивного фосфора 32Р и применения других тонких методик для изучения циркуляции фосфора в озерах. Хатчинсон (1957) и Пом-рой (1970) опубликовали обзоры работ по круговороту фосфора и других жизненно важных элементов.[ ...]
Эти работы показали, что, как правило, движение фосфора из организма в среду и обратно не происходит равномерно и постоянно, как можно подумать, глядя на схему (фиг. 34), хотя отмечалось, что на протяжении длительных периодов времени процессы в общем уравновешены. В любой момент времени большая часть фосфора находится в связанном состоянии — либо в организмах, либо в отложениях (в органическом детрите и неорганических частицах). Не более 10% фосфора присутствует в озерах в растворимой форме. Быстрое движение в обе -стороны (обмен) идет постоянно, но существенный обмен между твердой и растворимой формами часто нерегулярен, идет «рывками», с периодами, когда фосфор только уходит из отложений, и периодами, когда он только усваивается организмами или поступает в отложения, что связано с сезонными изменениями температуры и активности организмов. Как правило, связывание фосфора идет быстрее, чем освобождение. Растения быстро накапливают фосфор в темноте и в других условиях, когда не могут его использовать. За период быстрого роста продуцентов (обычно это бывает весной) весь доступный фосфор может оказаться связанным в продуцентах и консументах. Тогда активность -системы снижается, пока не разложатся трупы, фекалии и не высвободятся биогенные элементы. Однако концентрация фосфора в данный конкретный момент мало может сказать о продуктивности экосистемы. Низкое содержание растворенного фосфата может означать, либо что -система истощена, либо что метаболизм ее весьма интенсивен; только измерив скорость потока вещества, можно понять ситуацию. Помрой (1960) так формулирует это важное положение: «Измерение концентрации растворенного фосфата в природных водоемах не дает представления о доступности фосфора. Большая его часть или даже весь фосфор системы в любой момент может находиться в живых организмах, но при этом он может совершать полный «оборот» за один час, и в результате для организмов, способных поглощать фосфор из очень разбавленных растворов, его запас все время будет достаточным. Такие системы долгое время могут оставаться биологически стабильными при кажущемся отсутствии доступного фосфора. Изложенные здесь данные позволяют предположить, что быстрый поток фосфора типичен для высокопродуктивных систем и что для поддержания высокой продукции ■органики скорость потока важнее, чем концентрация элемента».[ ...]
Выше (гл. 2) подчеркивалось, что водоемы — не замкнутые системы; они должны рассматриваться как часть более обширных водосборных бассейнов. Как уже сказано, водосборная система — минимальная единица экосистемы, которой может практически управлять человек. На фиг. 37 показана количественная модель круговорота кальция для небольших гористых н лесистых водосборных бассейнов в Нью-Гемпшире площадью от 12 до 48 га (Борман и Лайкенс, 1967). Количество осадков (123 см в год) было измерено гидрометрическими станциями.[ ...]
Количество воды, поступаю щей в систему, измеряли у У-образной плотины(фиг.6).[ ...]
Определив концентрацию минеральных веществ в поступающей и оттекающей воде, можно рассчитать бюджет минеральных веществ. Содержание кальция в биотическом и почвенном фондах было оценено по данным Овингтона (1962).[ ...]
Механизмы удержания кальция и его возвращения в круговорот в ненарушенном лесу оказались столь эффективными, что, судя по полученным данным, экосистема теряла в год всего 8 кг кальция с гектара (и соответственно небольшие количества других биогенных элементов). Так как 3 кг кальция поступало в систему с дождем, то для установления равновесия достаточно было добавления 5 кг кальция на 1 га; такое количество кальция, как полагают, вполне может поступать из подстилающих пород (резервного фонда) за счет естественного выветривания. Недавняя работа Томаса (1969), в которой он для измерения скорости оборота использовал изотоп 45Са, показала, что деревья подлеска, например дёрен, работают как «кальциевые насосы», противодействующие движению кальция в глубь почвы и этим сохраняющие кальций в круговороте между организмами и активными верхними слоями подстилки и почвы.[ ...]
Числа обозначают количество кальция (в килограммах иа I га в год) В прямоугольниках приведены величины, характеризующие содержимое основных резервуа ров, или «неподвижных фондов».[ ...]
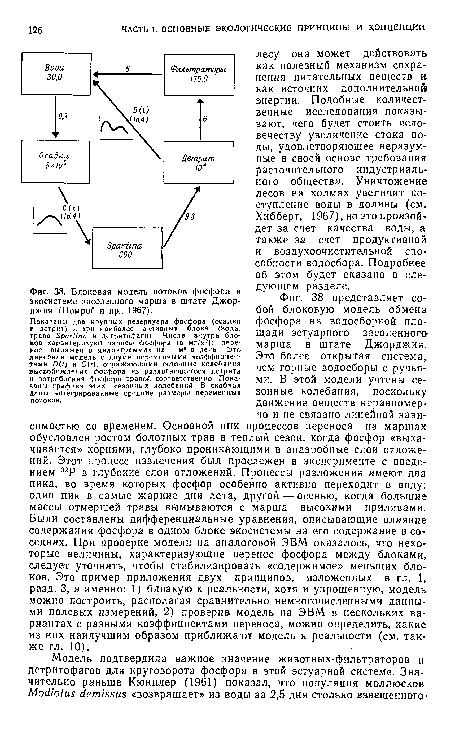
Показаны два крупных резервуара фосфора (осадки и детрит) и трн наиболее активных блока (вода, трава раг11па и детрнтофагн) Числа внутри блоков характеризуют запасы фосфора (в мг/м2); перенос выражен в миллиграммах на I м в день Это линейная модель с двумя переменными коэффициентами 0(0 и С(/), отражающими сезонные колебания высвобождения фосфора из разлагающегося детрита и потребления фосфора травой соответственно Показаны графики этих сезонных колебаний В скобках даны интегрированные средние размеры переменных потоков.[ ...]
Рисунки к данной главе:
| Блоковая модель потоков фосфора в экосистеме засоленного марша в штате Джорджия (Помрой и др., 1967). |
 |