Перенос энергии пищи от ее источника — растений —через ряд организмов, происходящий путем поедания одних организмов другими, называется пищевой цепью. При каждом очередном переносе большая часть (80—90%) потенциальной энергии теряется, переходя в тепло. Это ограничивает возможное число этапов, или «звеньев» цепи, обычно до четырех-пяти. Чем короче пищевая цепь (или чем ближе организм к ее началу), тем больше количество доступной энергии. Пищевые цепи можно разделить на два основных типа: пастбищные цепи, которые начинаются с зеленого растения и идут далее к пасущимся, растительноядным животным (т. е. к организмам, поедающим зеленые растения) и к хищникам (организмам, поедающим животных), и детритные цепи, которые начинаются от мертвого органического вещества, идут к микроорганизмам, которые им питаются, а затем детритофагам и к их хищникам. Пищевые цепи не изолированы одна от другой, а тесно переплетены. Их сплетение часто называют пищевой сетью. В сложном природном сообществе организмы, получающие свою пищу от растений через одинаковое число этапов, считаются принадлежащими к одному трофическому уровню. Так, зеленые растения занимают первый трофический уровень (уровень продуцентов), травоядные — второй (уровень первичных консументов), хищники, поедающие травоядных, — третий (уровень вторичных консументов), а вторичные хищники — четвертый уровень (уровень третичных консументов). Необходимо подчеркнуть, что эта трофическая классификация делит на группы не сами виды, а их типы жизнедеятельности; популяция одного вида может занимать один или более трофических уровней, смотря по тому, какие источники энергии она использует. Поток энергии через трофический уровень равен общей ассимиляции (Л) на этом уровне, а общая ассимиляция в свою очередь равна продукции биомассы (Р) плюс дыхание (/?) .[ ...]
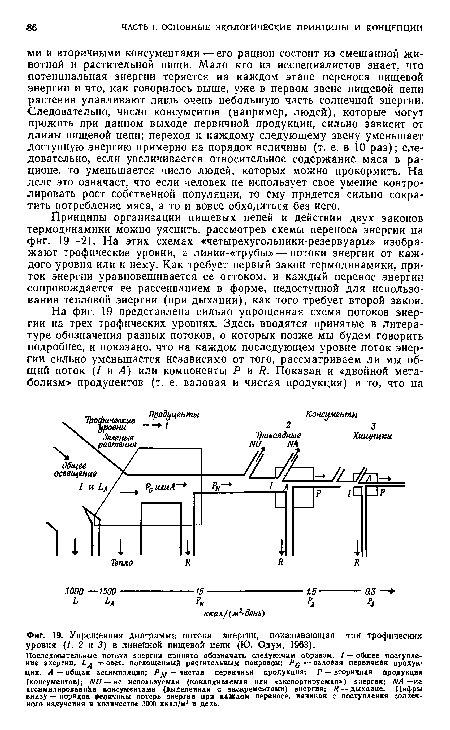
Принципы организации пищевых цепей и действия двух законов термодинамики можно уяснить, рассмотрев схемы переноса энергии на фиг. 19—21. На этих схемах «четырехугольники-резервуары» изображают трофические уровни, а линии-«трубы» — потоки энергии от каждого уровня или к нему. Как требует первый закон термодинамики, приток энергии уравновешивается ее оттоком, и каждый перенос энергии сопровождается ее рассеиванием в форме, недоступной для использования тепловой энергии (при дыхании), как того требует второй закон.[ ...]
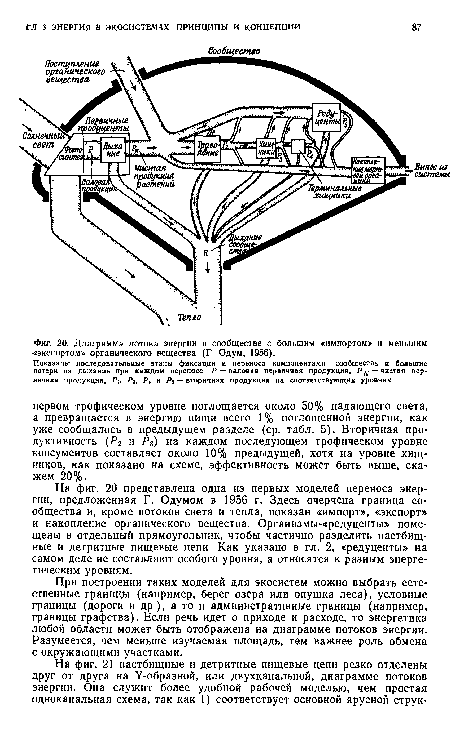
На фиг. 20 представлена одна из первых моделей переноса энергии, предложенная Г. Одумом в 1956 г. Здесь очерчена граница сообщества и, кроме потоков света и тепла, показан «импорт», «экспорт» и накопление органического вещества. Организмы-«редуценты» помещены в отдельный прямоугольник, чтобы частично разделить пастбищные и детритные пищевые цепи Как указано в гл. 2, «редуценты» на самом деле не составляют особого уровня, а относятся к разным энергетическим уровням.[ ...]
При построении таких моделей для экосистем можно выбрать естественные границы (например, берег озера или опушка леса), условные границы (дороги и др), а то и административные границы (например, границы графства). Если речь идет о приходе и расходе, то энергетика любой области может быть отображена на диаграмме потоков энергии. Разумеется, чем меньше изучаемая площадь, тем важнее роль обмена с окружающими участками.[ ...]
Такая гибкость в использовании модели может смутить начинающего. Необходимо еще раз подчеркнуть, что концепция трофического уровня введена не для того, чтобы классифицировать виды. Энергия течет через сообщество по этапам в соответствии со вторым законом термодинамики, но отдельная популяция вида может быть вовлечена не в один, а в несколько этапов, или трофических уровней (и очень часто именно так и бывает). Поэтому универсальную модель потока энергии, приведенную на фиг. 22, можно использовать двояко. Она может представлять популяцию вида — в этом случае каналы притока энергии и связи с другими видами составят обычную диаграмму пищевой сети с названиями отдельных видов в ее узлах (фиг. 23), или же может изображать энергетический уровень — тогда прямоугольники биомассы и каналы потоков энергии представляют все популяции (или части их), поддерживаемые одним источником энергии. Например, лисы обычно часть своей пищи получают от растений (плоды и т. д.), а часть их пищи составляют травоядные животные (зайцы, полевые мыши и т. д.). Если мы хотим подчеркнуть аспект внутрипопуляционной энергетики, то всю популяцию лис можно изобразить одним прямоугольником. Если же мы хотим распределить метаболизм популяции лис на два трофических уровня соответственно пропорции растительной и животной пищи, то надо изобразить два или несколько прямоугольников (как это сделано на фиг. 22, внизу справа). Так мы можем включить популяцию лис в общую схему потока энергии в сообществе. При изображении •целого сообщества нельзя смешивать эти два способа, кроме тех случаев, когда каждый вид занимает не более одного трофического уровня (например, сильно упрощенная экосистема «трава—корова—человек»).[ ...]
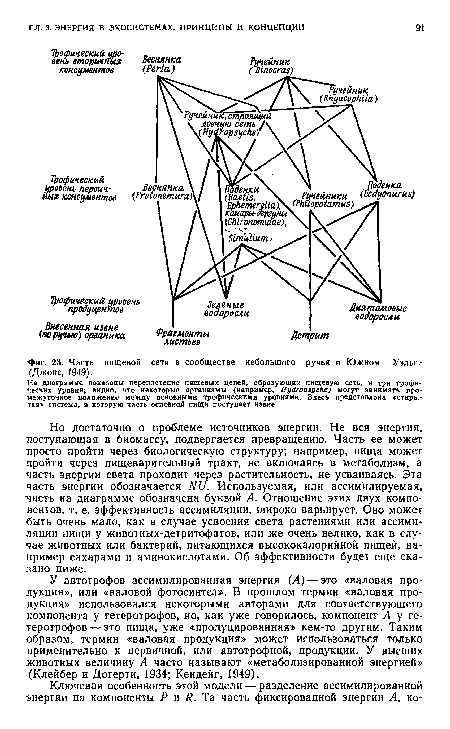
На диаграмме показаны переплетение пищевых цепей, образующих пищевую сеть, и три трофических уровня; видно, что некоторые организмы (например, Нуйторьуске) могут занимать промежуточное положение между основными трофическими уровнями. Здесь представлена «открытая» система, в которую часть основной пищи поступает извне.[ ...]
Но достаточно о проблеме источников энергии. Не вся энергия, поступающая в биомассу, подвергается превращению. Часть ее может просто пройти через биологическую структуру; например, пища может пройти через пищеварительный тракт, не включаясь в метаболизм, а часть энергии света проходит через растительность, не усваиваясь. Эта часть энергии обозначается N1/. Используемая, или ассимилируемая, часть на диаграмме обозначена буквой А. Отношение этих двух компонентов, т. е. эффективность ассимиляции, широко варьирует. Оно может ■быть очень мало, как в случае усвоения света растениями или ассимиляции пищи у животных-детритофагов, или же очень велико, как в случае животных или бактерий, питающихся высококалорийной пищей, например сахарами и аминокислотами. Об эффективности будет еще сказано ниже.[ ...]
У автотрофов ассимилированная энергия (А) — это «валовая продукция», или «валовой фотосинтез». В прошлом термин «валовая продукция» использовался некоторыми авторами для соответствующего компонента у гетеротрофов, но, как уже говорилось, компонент А у ге-теротрофов — это пища, уже «продуцированная» кем-то другим. Таким образом, термин «валовая продукция» может использоваться только применительно к первичной, или автотрофной, продукции. У высших животных величину А часто называют «метаболизированной энергией» (Клейбер и Догерти, 1934; Кендейг, 1949).[ ...]
Рисунки к данной главе:
| Упрощенная диаграмма потока энергии, показывающая три tpoфичecкиx уровня (1, 2 н 3) в линейной пищевой цепи (Ю. Одум, 1963). |
 |
| Диаграмма потока энергии в сообществе с большим «импортом» и меньшим -«экспортом» органического вещества (Г Одум, 1956). |
 |
| Компоненты «универсальной» модели экологического потока энергии (Ю. Одум, 1968). |
 |
| Часть пищевой сети в сообществе небольшого ручья в Южном Уэльса (Джонс, 1949). |
 |
| Схема основных пищевых цепей в пруду (штат Джорджия) для спортивной рыбиой ловли. |
 |
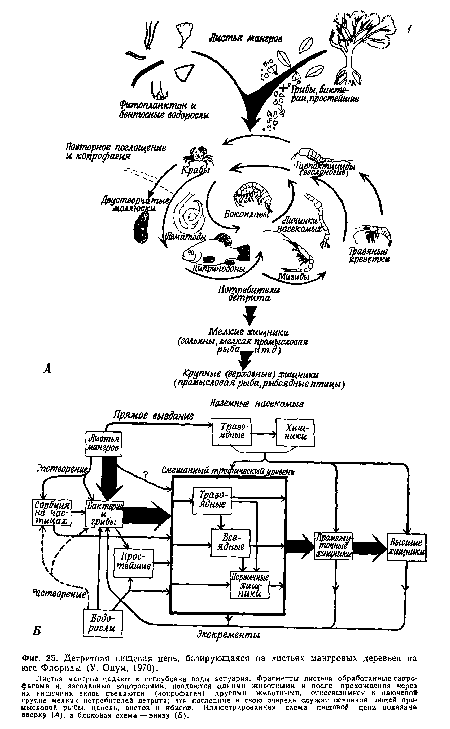
| Детритная пищевая цепь, базирующаяся на листьях мангровых деревьев на юге Флориды (У. Одум, 1970). |
 |
| Связь между эффективностью роста тканей, экологической эффективностью роста и эффективностью ассимиляции в популяциях животных (Уэлч, 1968). |
 |