Другую интересную группу, известную как хемосинтезирующие бактерии, часто рассматривают как «продуцентов» (хемоавтотрофов), однако с точки зрения их роли в экосистемах это группа, переходная между автотрофами и гетеротрофами. Хемосинтезирующие бактерии получают энергию, необходимую для включения двуокиси углерода в состав компонентов клетки, не за счет фотосинтеза, а за счет химического окисления таких простых неорганических соединений, как аммоний (окисляется в нитрит), нитрит (в нитрат), сульфид (в серу), закись железа (в окись). Они могут расти в темноте, но большинство нуждается в кислороде. Представители этой группы — серная бактерия Яе 1‘а/оа, встречающаяся в изобилии в серных источниках, и различные азотные бактерии, играющие важную роль в круговороте азота (гл. 4, разд. 1). Одна из уникальных групп хемосинтезирующих бактерий— водородные бактерии — серьезно рассматривается как возможный компонент систем поддержания жизни в космических кораблях, так как они могут очень эффективно удалять углекислоту из атмосферы корабля (см. гл. 20, разд. 1). Благодаря способности существовать в темноте (в почве и донных отложениях) хемосинтезирующие бактерии не только играют роль в извлечении минеральных питательных веществ, но, как показал русский гидробиолог И. Т. Сорокин (1966), осваивают энергию, которая иначе была бы потеряна для животных.[ ...]
Процесс производства пищи путем фотосинтеза часто называют «работой зеленых растений». Как теперь полагают, синтез аминокислот, белков и других жизненно важных веществ происходит одновременно с синтезом углеводов (глюкозы) и основывается на тех же главных реакциях. Часть синтезируемой пищи используется, разумеется, самими продуцентами. Излишек, равио как и протоплазма продуцентов, используется затем консументами или, как отмечалось, частично накапливается или переносится в другие системы. Динамика «продуцирования» будет подробно рассмотрена в гл. 3.[ ...]
Аэробное дыхание — процесс, обратный «нормальному» фотосинтезу (см. словесную формулу фотосинтеза, приведенную выше). С помощью этого процесса все высшие растения и животные, а также большинство бактерий и простейших получают энергию для поддержания жизнедеятельности и построения клеток. В итоге завершенного дыха-, ния образуются СОг, Н2О и вещества клетки, однако процесс может идти не до конца и в результате такого незавершенного дыхания образуются органические вещества, еще содержащие некоторое количество энергии, которая может быть в дальнейшем использована другими организмами.[ ...]
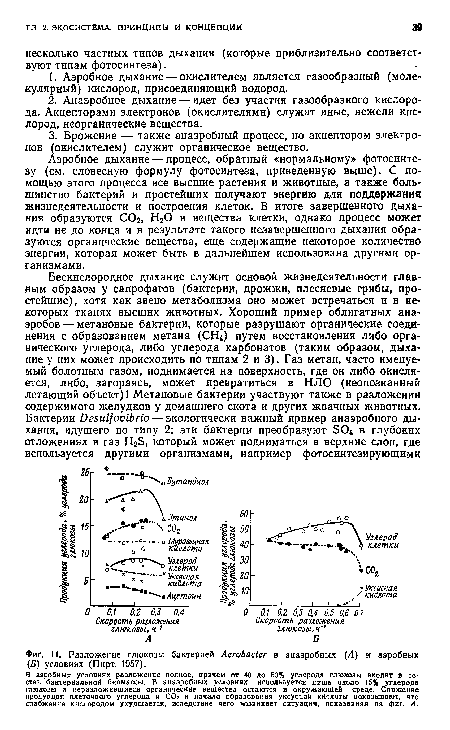
Как уже указывалось, многие группы бактерий способны й к аэробному, и к анаэробному дыханию (т. е. являются факультативными анаэробами), но важно отметить, что конечные продукты этих двух реакций различны и количество высвобождающейся энергии в анаэробных условиях значительно меньше. На фиг. 11 представлены результаты интересного исследования, в котором один и тот же вид бактерий, АегоЬаЫег, выращивали в анаэробных и аэробных условиях с использованием глюкозы в качестве источника углерода. В присутствии кислорода почти вся глюкоза превращалась в бактериальную протоплазму и СО2, в отсутствие же кислорода разложение было неполным, гораздо меиьшая часть глюкозы превращалась в вещество клетки, и в среду выделялся ряд органических соединений, для окисления которых требуются дополнительные «специалисты»-бактерии. В общем полное аэробное дыхание во много раз быстрее, чем неполный процесс анаэробного дыхания, если оценивать выход энергии на единицу используемого субстрата.[ ...]
Когда скорость поступления органического детрита в почву и донные отложения высока, бактерии, грибы, простейшие и другие организмы создают анаэробные условия, используя кислород быстрее, чем он может диффундировать в субстрат; разложение органики при этом не прекращается — оно продолжается, хотя и с меньшей скоростью, если среди микробного населения представлены организмы с соответствующими типами анаэробного метаболизма.[ ...]
Итак, несмотря на то что анаэробные сапрофаги, как облигатные, так и факультативные, составляют меньшую часть компонентов сообщества, они тем не менее играют в экосистеме важную роль, так как лишь они способны к дыханию в лишенных света бескислородных нижних ярусах системы. Занимая эти негостеприимные местообитания, они «спасают» энергию и материалы, делая их доступными для большинства аэробов. Таким образом, то, что кажется «неэффективным» способом дыхания, оказывается составной частью «эффективной» эксплуатации энергии и материальных ресурсов экосистемой в целом. Например, эффективность очистки сточных вод, которая обеспечивается управляемой человеком гетеротрофной экосистемой, зависит от согласованности между деятельностью анаэробных и аэробных сапрофагов. Общая картина взаимодействия анаэробов и аэробов в осадочных образованиях показана на фиг. 35 и 166.[ ...]
Как было подчеркнуто в разд. «Определения», для биосферы в целом важнейшее значение имеет отношение темпов продуцирования и разложения независимо от того, какие организмы или абиотические процессы определяют оба эти процесса. Соотношение этих противоположных функций контролирует состав атмосферы и гидросферы, и, как уже подчеркивалось, к счастью для человека с его огромными машинами, потребляющими кислород, продуцирование до сего времени преобладало над разложением. Однако сейчас человек «берет больше, нежели отдает», и это несоответствие возросло настолько, что угрожает нарушением жизненно важного баланса в природе. Отставание гетеротрофной утилизации продуктов автотрофного метаболизма есть, следовательно, одно из наиболее важных свойств экосистемы, и оно находится под угрозой из-за беспечности человека. Имеет смысл рассмотреть здесь основные особенности процесса разложения, несмотря на то, что многие детали будут обсуждаться далее.[ ...]
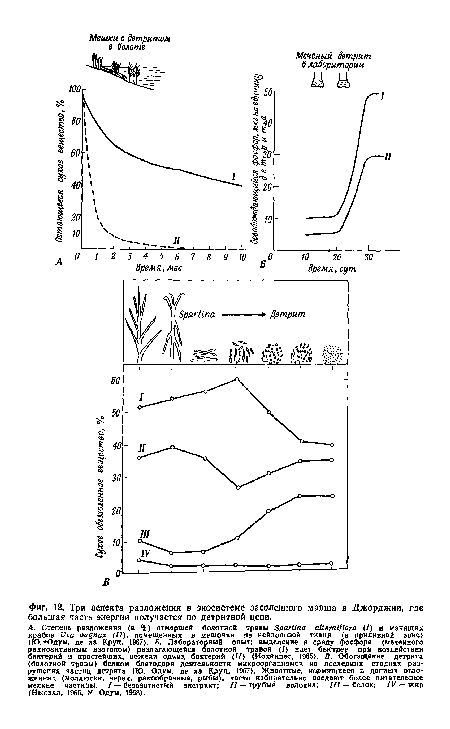
Разложение включает как абиотические, так и биотические процессы. Так, например, степные и лесные пожары — это не только важные лимитирующие факторы (см. гл. 5), но также «агенты разложения» детрита, возвращающие большое количество СОг и других газов в атмосферу и минеральных веществ в почву. Однако обычно мертвые растения и животные разлагаются гетеротрофными микроорганизмами и сапрофагами. Такое разложение есть способ, посредством которого бактерии и грибы получают для себя пищу. Разложение, следовательно, происходит благодаря энергетическим превращениям в организмах и между ними. Этот процесс абсолютно необходим для жизни, так как без него все питательные вещества оказались бы связанными в мертвых телах и никакая новая жизнь не могла бы возникать. В бактериальных клетках и мицелии грибов имеются наборы ферментов, необходимых для осуществления специфических химических реакций. Эти ферменты выделяются в мертвое вещество; некоторые из продуктов его разложения поглощаются разлагающими организмами, для которых они служат пищей, другие остаются в среде; кроме того, некоторые продукты выводятся из клеток. Ни один вид сапротрофов не может осуществить полное разложение мертвого тела. Однако гетеротрофное население биосферы состоит из большого числа видов, которые, действуя совместно, производят полное разложение. Различные части растений и животных разрушаются с неодинаковой скоростью. Жиры, сахара и белки разлагаются быстро, а целлюлоза и лигнин растений, хитин, волосы и кости животных разрушаются очень медленно. Это показано на фиг. 12; здесь сравнивается интенсивность разложения мертвой массы болотных трав и манящих крабов (помещенных в нейлоновые мешочки) в засоленном марше в штате Джорджия. Отметим, что около 25% сухого веса трав разложилось за месяц, а остальные 75% разлагались медленнее. Через 10 мес. еще оставалось 40% первоначальной массы трав. Остатки же крабов исчезли к этому времени полностью. По мере того как детрит разрушается и удаляется из мешочка, оставшаяся часть в результате интенсивной деятельности микроорганизмов обогащается белком (фиг. 12, В), благодаря чему становится более питательной пища для детритоядных животных (Одум, де ла Круц, 1967; Кауши, Хайнес, 1968).[ ...]
Рисунки к данной главе:
| Разложение глюкозы бактерией АегоЬаЫег в анаэробных (А) и аэробных (Б) условиях (Пирт, 1957). |
 |
| Три аспекта разложения в экосистеме засоленного марша в Джорджии, где большая часть энергии получается по детритной цепи. |
 |
Аналогичные главы в дргуих документах:
| См. далее:Продуцирование и разложение в природе |