| Схема митотического цикла. После митоза (М) дочерние клетки вступают в фазу к концу которой происходит подготовка к синтезу ДИК. Синтез ДНК и гистонов происходит в следующей Б-фазе; в это же время происходит репликация хромосом, в результате чего содержание ДНК в клетке удваивается. Затем клетка вступает в фазу С2, представляющую собой период подготовки к митозу, включающий конденсацию хромосом. |
 |
Далее
| Образование клеточной пластинки путем слияния телец Гольджи. |
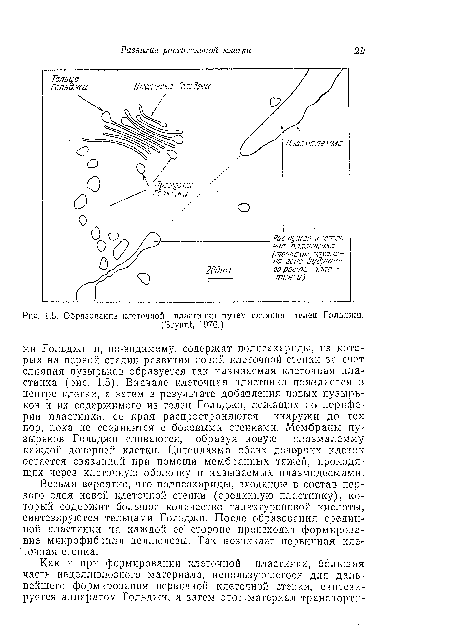 |
Далее
| Схема, иллюстрирующая отношение между эндоплазматнческим |
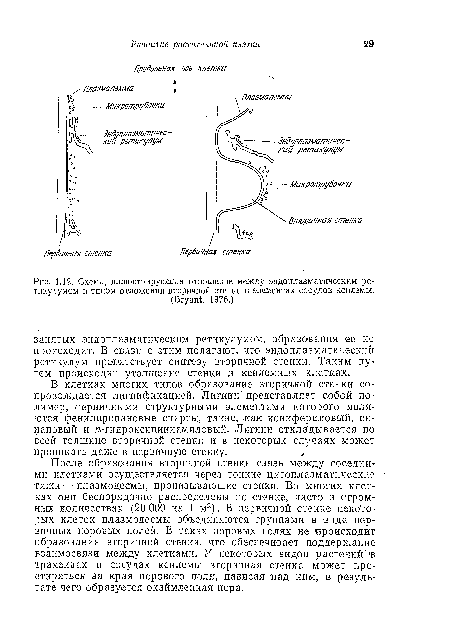 |
Далее
| Л. Неэквивалентное деление при формировании замыкающих клеток устьица в листе однодольного растения. После неэквивалентного деления эпидермальных клеток образуются меньшая устьнчиая материнская клетка (черное ядро) и большая клетка (белое ядро). Хлоропласта развиваются в устьпч-иой материнской клетке, которая вновь делится, причем плоскость этого деления располагается под прямым углом к плоскости первого деления, Образующиеся в результате дочерние клетки развиваются в замыкающие клетки устьица. Б, В. Развитие пыльцевого зерна. |
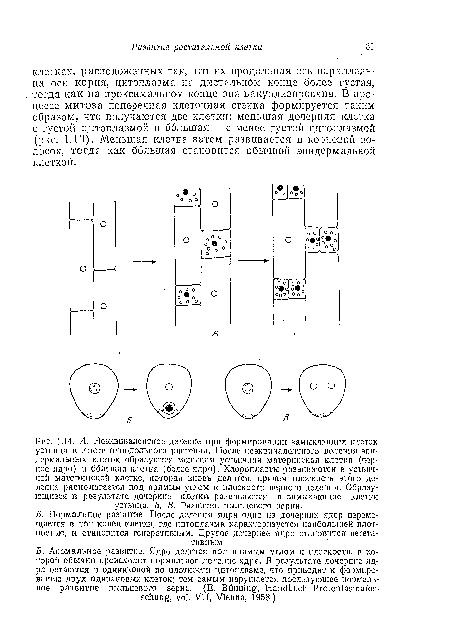 |
Далее
| Последовательные стадии раннего развития зародыша у Fucus. |
 |
Далее
| Радиоавтография среза кончика корня горчицы (Sinapis), сфотографированного в темном поле. (Clowes, Juniper, 1968.) |
 |
Далее
| Рост колонии дрожжей в постоянном объеме культуральной среды. |
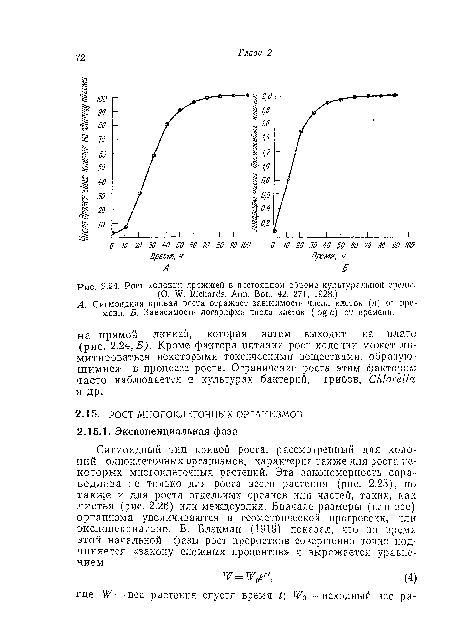 |
Далее
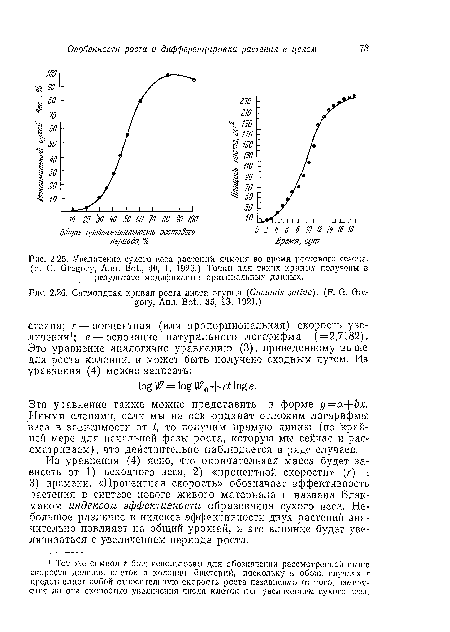
| Увеличение сухого веса растений ячменя во время ростового сезона. {F. G. Gregory, Ann. Bot., 40, 1, 1926.) Точки для таких кривых получены в результате модификации оригинальных данных. |
 |
Далее
| Сигмоидная кривая роста листа огурца (Cucumis saliva). (F. G. Gregory, Ann. Bot., 35, 93, 1921.) |
|
Далее
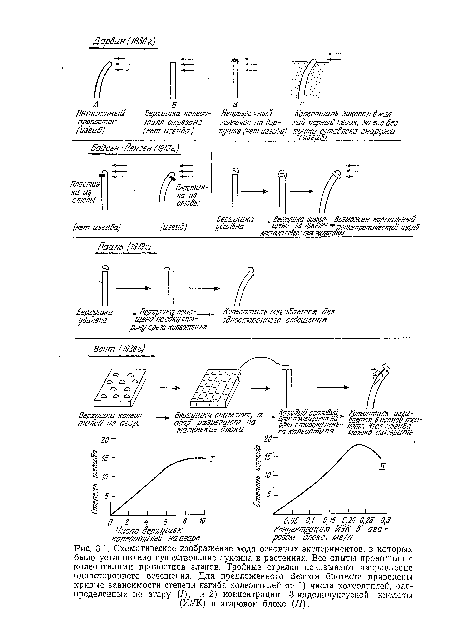
| Схематическое изображение хода основных экспериментов, в которых было установлено существование ауксина в растениях. Все опыты проводили с колеоптилями проростков злаков. Тройные стрелки показывают направление одностороннего освещения. Для предложенного Веитом биотеста приведены кривые зависимости степени изгиба колеоптилей от 1) числа колеоптилей, распределенных по агару (/), и 2) концентрации 3-индолилуксусной кислоты (ИУК) в агаровом блоке (II). |
 |
Далее
| Пути окисления ИУК. |
 |
Далее
| Известные способы биохимической инактивации абсцизовой кислоты |
 |
Далее
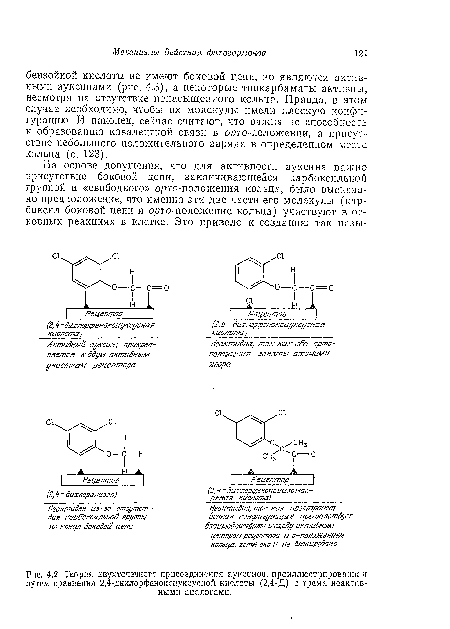
| Теория двухточечного присоединения ауксинов, проиллюстрированная путем сравнения 2,4-дихлорфеноксиуксусиой кислоты (2,4-Д) с тремя неактивными аналогами. |
 |
Далее
| Примеры различных молекул активных ауксинов. Их общей чертой является наличие сильного отрицательного заряда (—), отделенного от более слабого положительного заряда (6 + ) расстоянием в 5,5 А. (K. V. Thimann. Ann. Rev. Plant Physiol., 14, 1—18, 1963.) |
 |
Далее
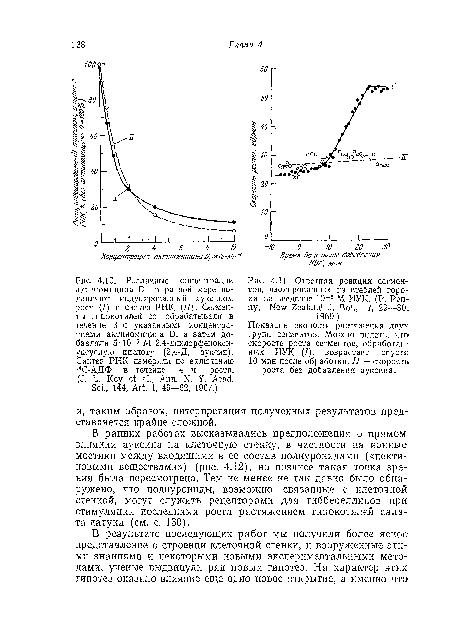
| Ответная реакция сегментов, изолированных из стеблей гороха, на введение 10“5 М ИУК. (P. Penny, New Zealand J. Bot., 7, 29—30,. |
 |
Далее
| Образование и взаимопревращение некоторых пектиновых веществ (полигалактуропидов) в стенках растительных клеток. |
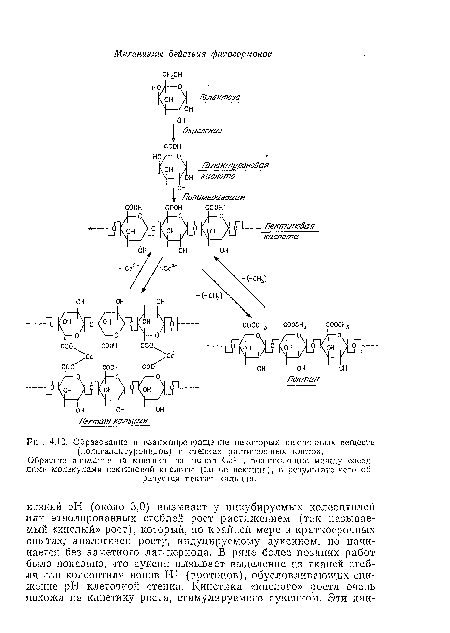 |
Далее
| Схема, иллюстрирующая гипотетическую модель влияния аукснпа (ИУК) на связанный с мембраной протонный (Н+) насос. (По P. J. Davis, The Botanical Review, 39, 139—171, 1973.) |
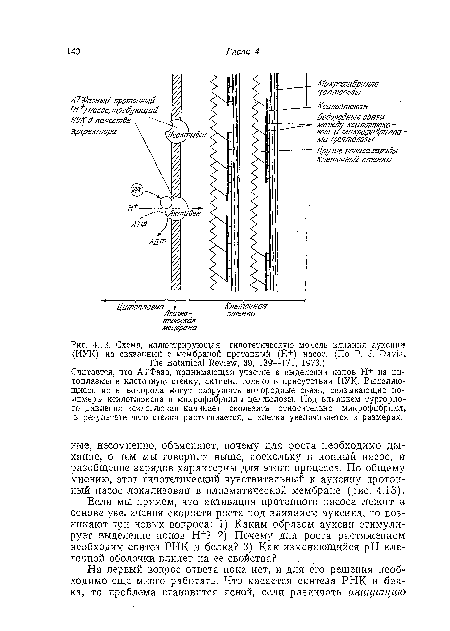 |
Далее
| Формула сериновой тРНК (выделенной из клеток дрожжей). |
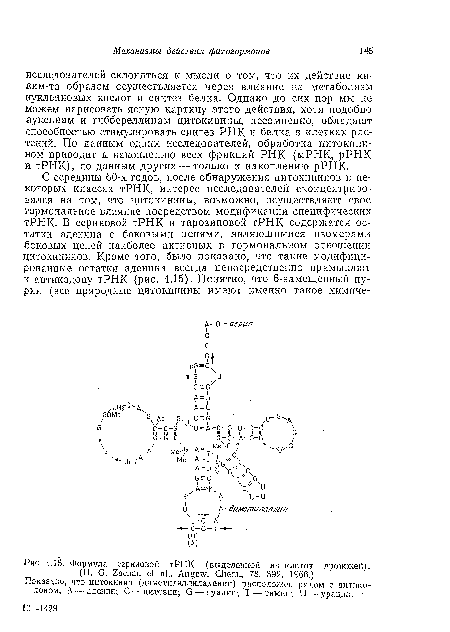 |
Далее
| Зависимость от времени высвобождения фермента (а-амилазы) из изолированных алейроновых слоев ячменя, инкубируемых на среде, содержащей 10 |
 |
Далее
| Влияние удаления и вторичного добавления гиббереллина (ГАз) к алейроновым слоям ячменя на синтез в1 них сс-амилазы. |
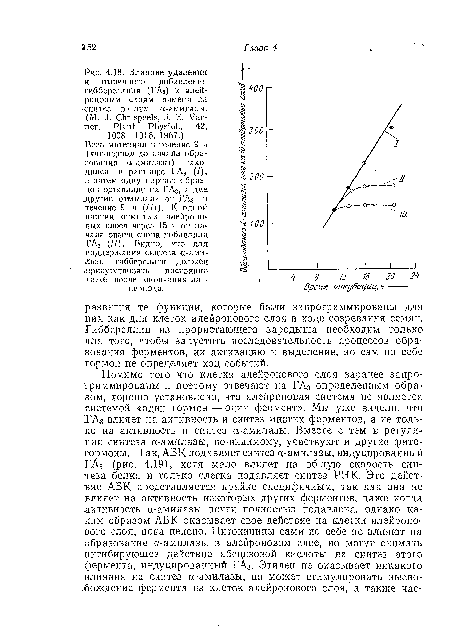 |
Далее
| Определение скорости ба-зипетального полярного транспорта 3-индолилуксусной кислоты (ИУК) в отрезках черешков фасоли. (С. С. McCready, W. P. Jacobs, |
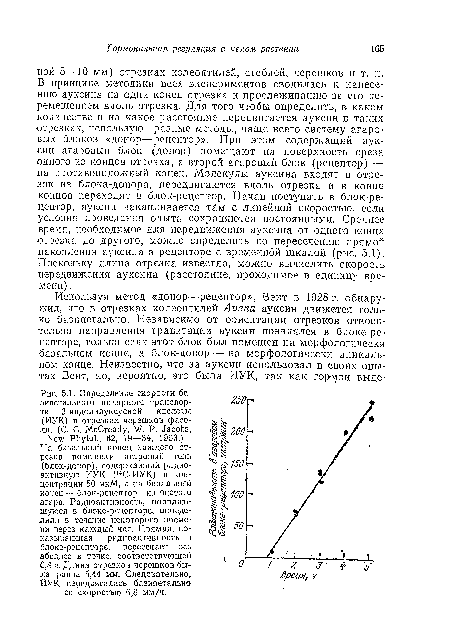 |
Далее
| А. Корреляция между скоростью роста (/) и содержанием ауксина (II) вдоль оси стебля проростка гороха (эпикотиль). Вверху |
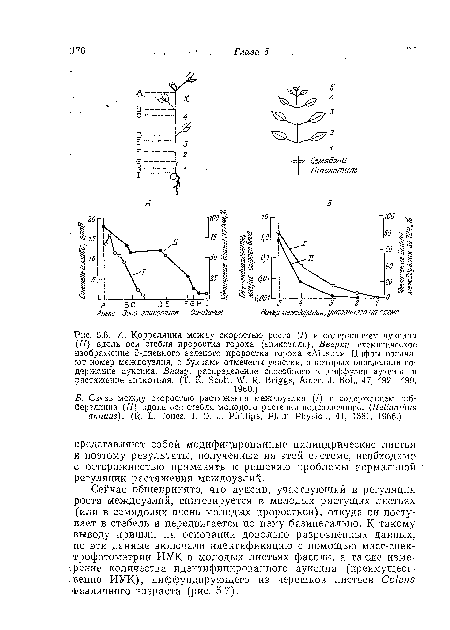 |
Далее
| А. Типичная концентрационная кривая влияния ауксина (3-иидолил- |
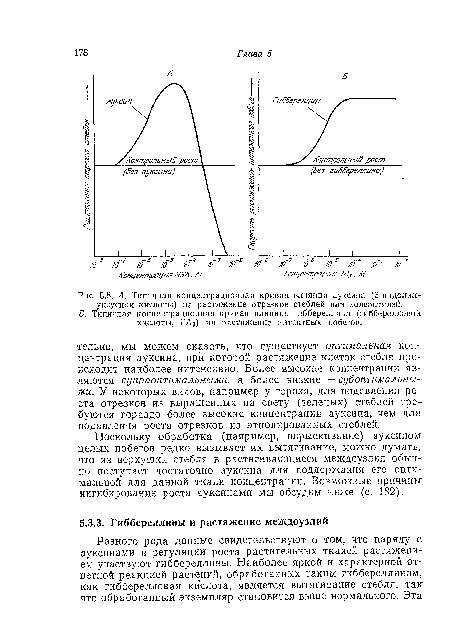 |
Далее
| Подавление этиленом растяжения |
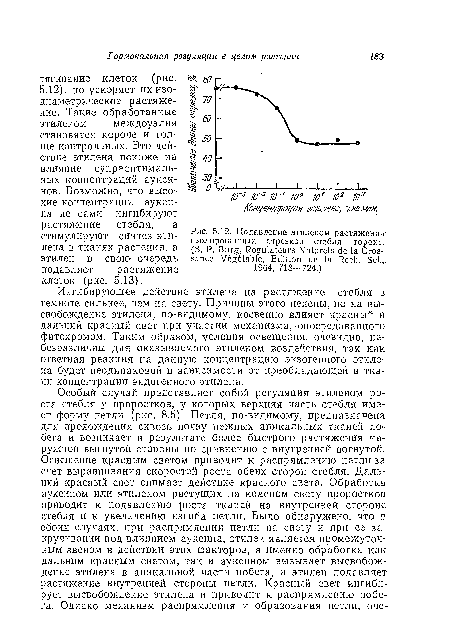 |
Далее
| М, а при более высоких концентрациях 2,4-Д скорость образования этилена сильно увеличивалась, а скорость растяжения уменьшалась. Создается впечатление, что у двудольных растений подавление роста высокими концентрациями ауксина опосредовано, их действием на синтез этилена и ингибиторным влиянием последнего. У однодольных может наблюдаться другая ситуация. |
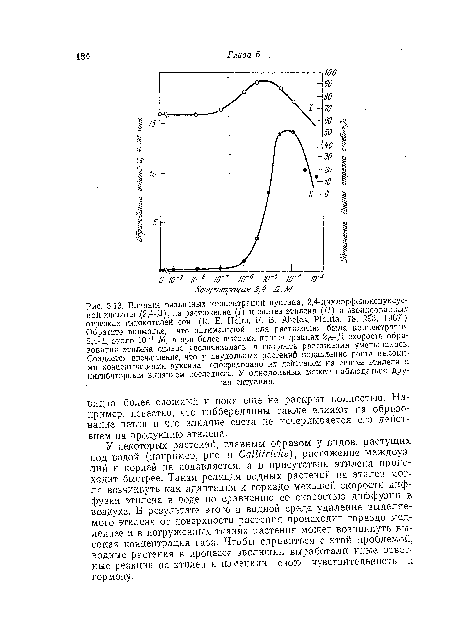 |
Далее
| Дифференцировка проводящей ткани в стерильной культуре каллуса. А. Дифференцировка проводящей ткани в каллусе вызвана присутствием привитой почки того же вида. (По Wetmore, Sorokin, 1955.) |
 |
Далее
| Индуцированное ауксином образование ксилемы га корне гороха. |
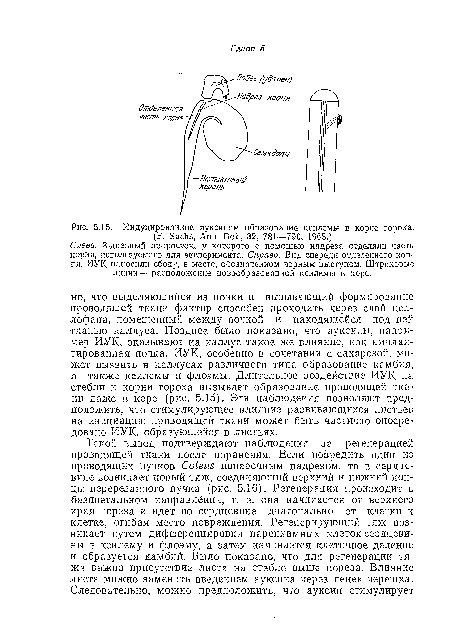 |
Далее
| Влияние гормонов на рост боковых почек после дека-питации растений гороха (Pi-sum sativum). (Данные |
 |
Далее
| Апикальное доминирование у растений фасоли (Р1гавео1и&). Шестнадцати дневные растения декапитировали за пять дней до того, как была сделана фотография. Слева направо |
 |
Далее
| Содержание в листьях Хапшит st/ пит разного возраста 1 4 |
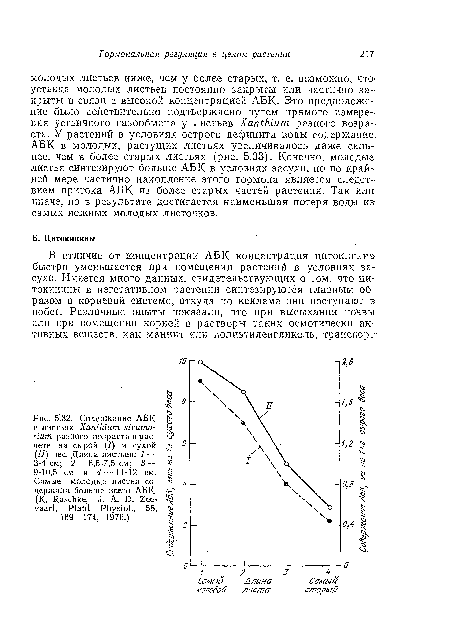 |
Далее
| Влияние опрыскивания гибберелловой кислотой (ГА3) па урожай сельдерея. (Фотография любезно предоставлена д-ром |
 |
Далее
| Эпинастические реакции корней растений томата па переувлажнение почвы (В), высокую концентрацию атмосферного этилена (В) или недостаточную аэрацию корневой системы, находящейся в питательном растворе (Г). (Фотографии любезно предоставлены д-ром М. В. Jackson, Великобритания.) |
 |
Далее
| Замена ауксином (ИУК) контактного раздражителя в индукции скручивания усика. (Фотография любезно предоставлена д-ром Leonora Reinhold, |
 |
Далее
| Спектр действия первой положительной реакции колеоптилей овса. Максимум кривой наблюдается при одностороннем освещении синим светом. (K. V. Thimann, G. М. Curry, Comparative Biochem., 1, 243—306, Academic |
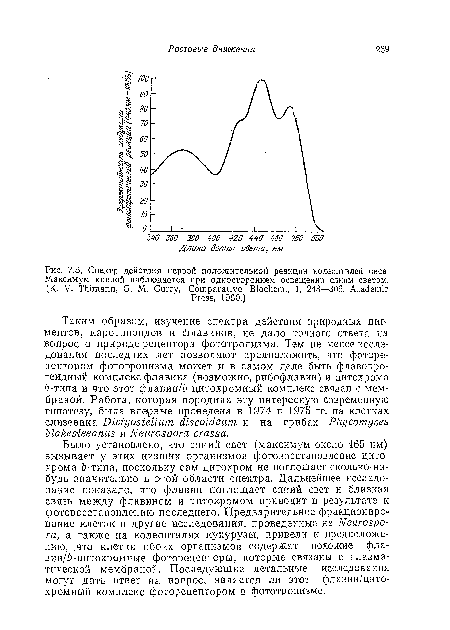 |
Далее
| Спектры поглощения рибофлавина (/) и |р-каротина (II). Ни один из них не совпадает со спектром действия фототропизма (рис. 7.5), одиако когда определение рибофлавина ведется в липоидном растворителе,^его спектр поглощения в большей степени сходен со спектром действия. |
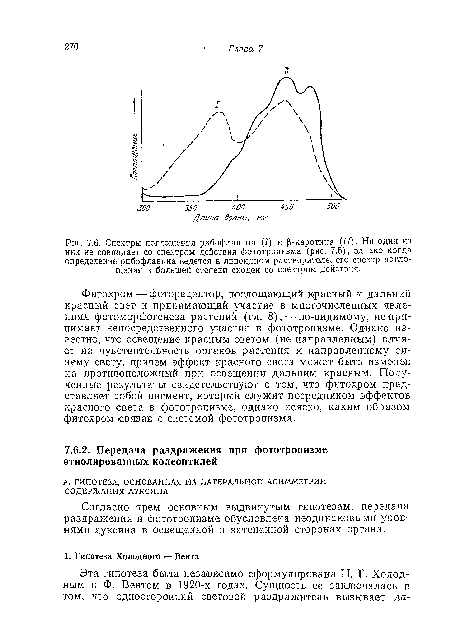 |
Далее
| Оригинальный эксперимент Beirr а, показавший, что при одностороннем освещении верхушек колеоптилей овса больше ауксина диффундирует в агаровый блок, расположенный под теневой стороной, чем в блок под освещенной стороной (Б). Общее количество ауксина, поступавшего из освещенных верхушек, было на 16% меньше, чем из верхушек, содержавшихся в темноте (Л). Данное различие, по-видимому, пе имеет существенного значения. |
 |
Далее
| Диффузия ауксина в агаровые блоки, происходящая из верхушек колеоптилей кукурузы, подвергнутых различным воздействиям. (W. R, Briggs, Я. D. Tocher, J. F. Wilson, Science, 126, 210—212, 1957.) |
 |
Далее
| Спектры действия регулируемых фитохромом физиологических реакций, возникающих на красный и дальний красный свет. Эффект красного света— сплошная линия, эффект дальнего красного — штриховая линия. (По F. В. Salisbury, Endeavour, 24, 78—80, 1965.) |
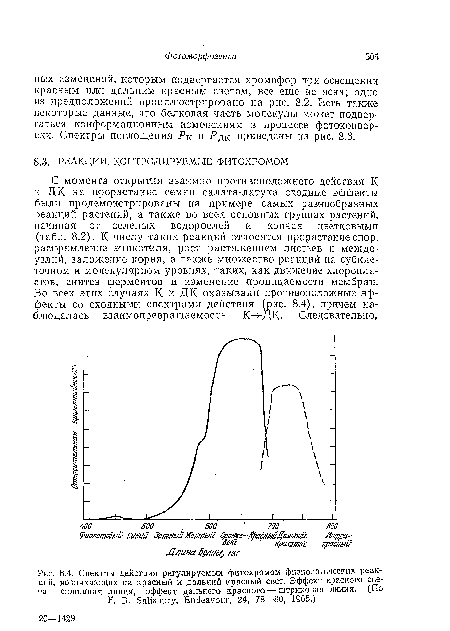 |
Далее
| Типичное спектральное распределение естественного дневного света до (/) и после (//) прохождения через полог растительности. (М. G. Holmes, |
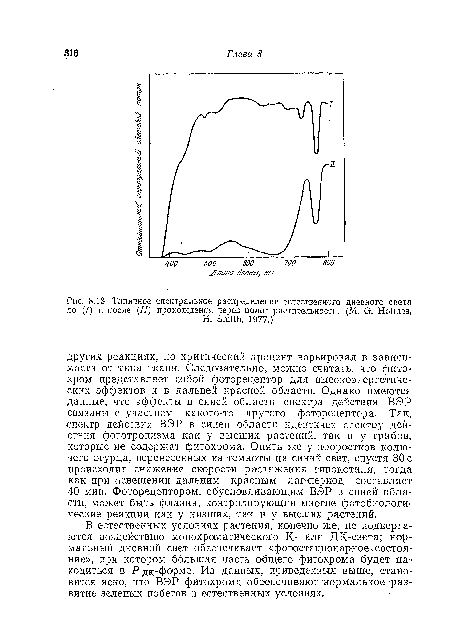 |
Далее
| Зависимость между равновесием ¿штохрома (ф) и коэффициентом квантового потока (£) при различных световых режимах. (М. G. Holmes, Н. Smith, 1977.) |
 |
Далее
| Зацветание хризантемы Происходит, если листья находятся в условиях короткого дня (Б, Г), независимо от того, находятся ли апексы в условиях длинного (5) или.короткого (Г) дня. " |
 |
Далее
| Влияние изменения длины фотопериода при постоянном темновом периоде продолжительностью 16 ч на цветение сои. (К. С. Hamner, Bot. Gaz., |
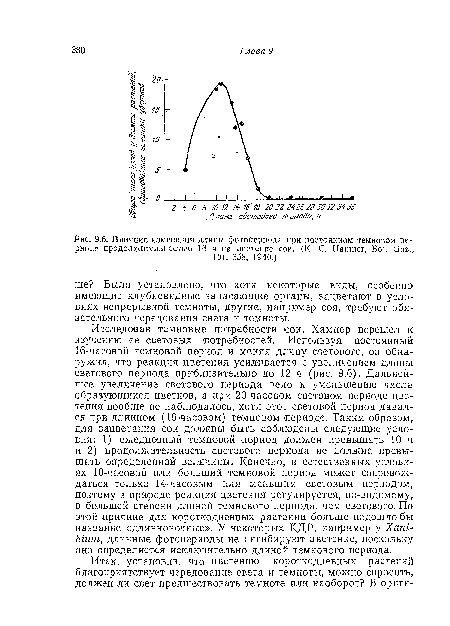 |
Далее
| Гипотетические изменения количества фитохрома .Рдк в листьях во время темпового периода, прерванного кратковременным действием красного |
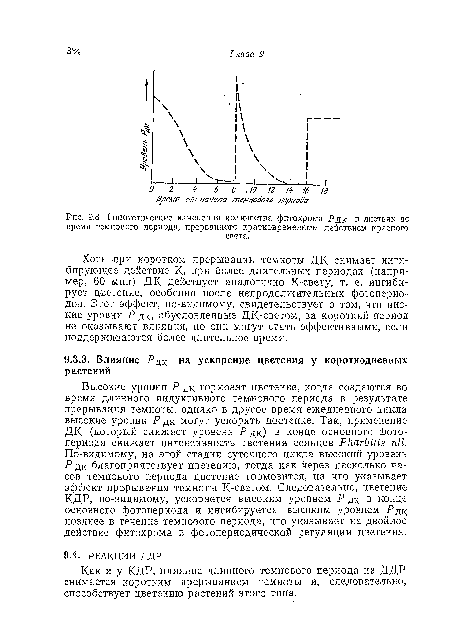 |
Далее
| Реакция короткодиевных и длиннодиевных растений при различных фотопериодических режимах. |
 |
Далее
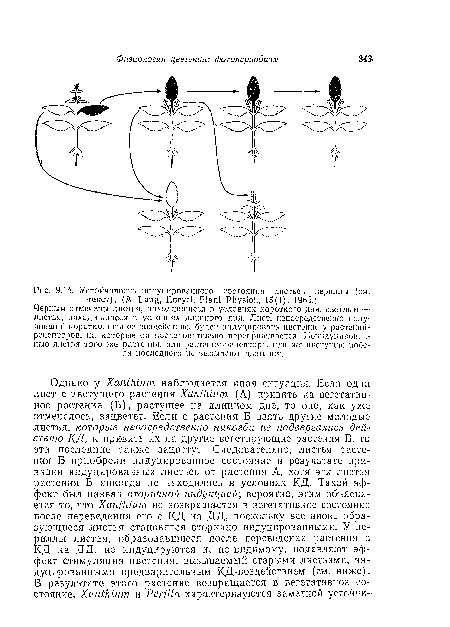
| Вверху. Растения Xanihium, имеющие две ветви. Одна из ветвей находилась в условиях короткого дня, а остальная часть растения находилась в условиях длшшого дня. Оба растения цвели, если побег, находившийся на коротком дне, имел по меньшей мере >/в листа. При полном удалении листьев цветение не наступало (справа). |
 |
Далее
| Устойчивость индуцированного состояния листьев периллы (см. |
 |
Далее
| Реакция цветения фасоли Билокси на циклы различной длины. (Eta К- Hamner, Environmental Control of Plant Growth, Academic Press, New York, |
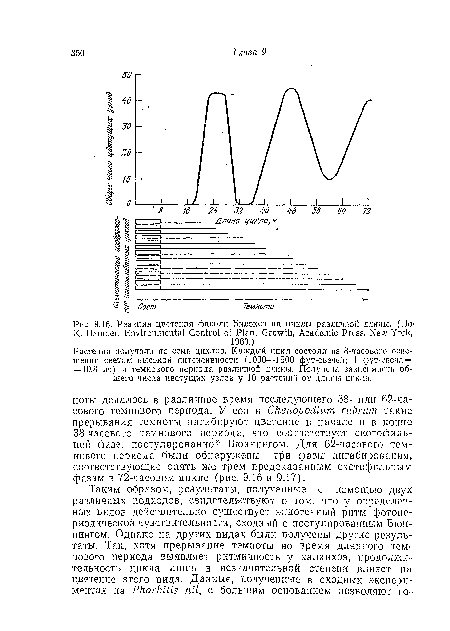 |
Далее
| Влияние яровизации и гпбберелловой кислоты на цветение моркови. |
 |
Далее
| Черепки, взятые от «взрослой» части плюща (Hederá helix), в течение нескольких лет продолжают сохранять «взрослые» признаки, такие, как форма листа, филлотаксис и способность к цветению, хотя высокие температуры способствуют возврату к ювенильному состоянию. (Напечатано с разрешения д-ра L. W. Robinson.) |
 |
Далее
| Влияние послеуборочного дозревания при сухом хранении в условиях комнатной температуры на скорость прорастания семян Impaiiens balsamina. |
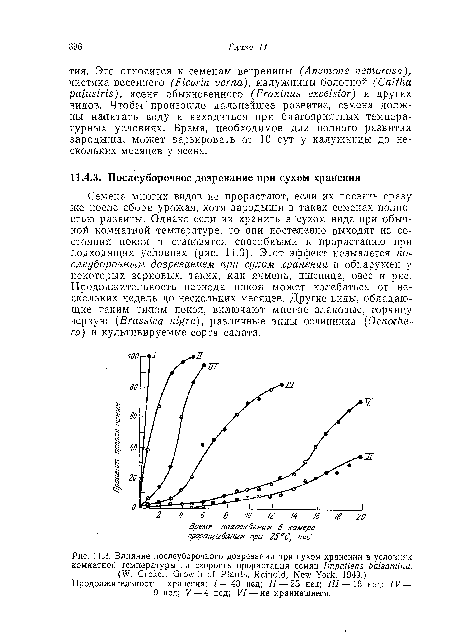 |
Далее
| Типы старения растений и листьев. |
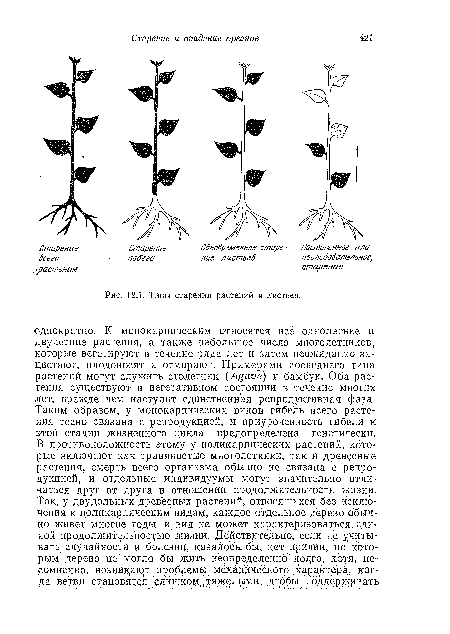 |
Далее
| Изменение в содержании белкового и растворимого азота в отделенных листьях табака. После срезания листа количество белкового азота уменьшает- ^ ся, а количество растворимого азота увеличивается. |
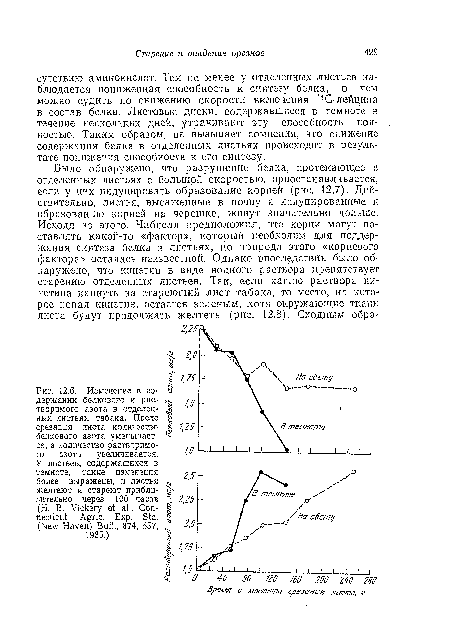 |
Далее
| Эффект действия кинетина па старение листа. |
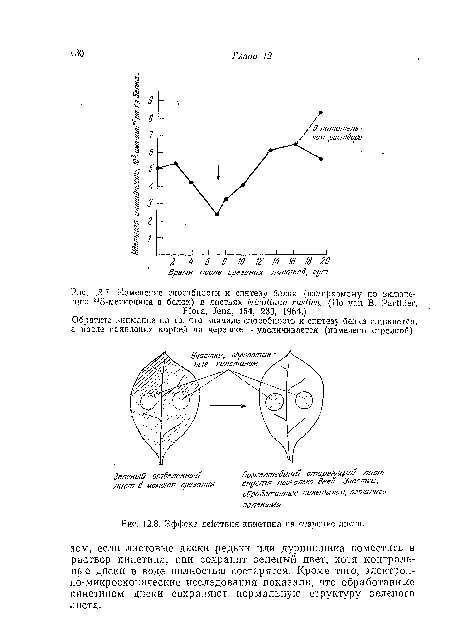 |
Далее
| Продольный срез зоны старения в основании черешка листа типичного двудольного растения. (J. Torrey, Development in Flowering Plans, Macmillan, New York, 1967.) |
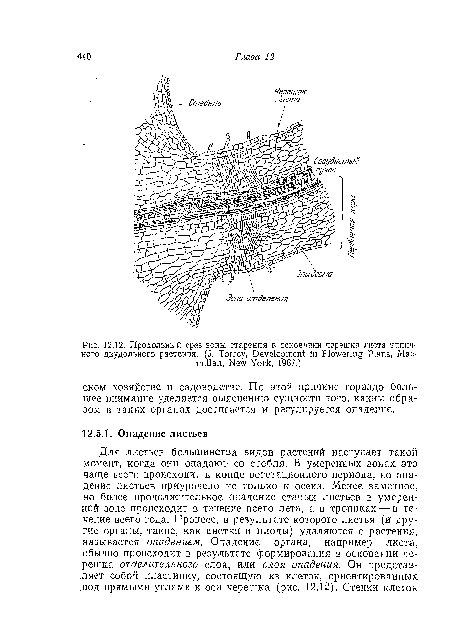 |
Далее
| Три типа расположения повторяющейся ДНК, характерные для хромосом высших растений. (Из R. Flavell, 1980.) |
 |
Далее
| Схема синтеза и процессинга предшественника рибосомной РНК и |
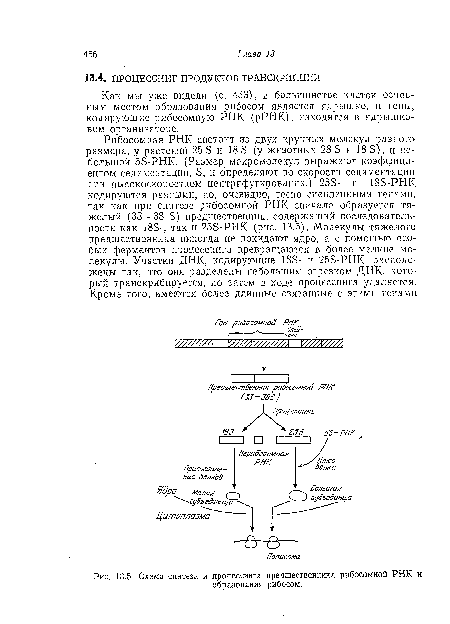 |
Далее
| Схема, иллюстрирующая теорию регуляции синтеза белка Жакоба |
 |
Далее
| А. Взрослые растения Acetabularia mediterránea. Б. Срез через шапочку, на котором видно образование цист. |
 |
Далее
| Схема по снятию состояния привыкания. (F. Meins, 1977.) |
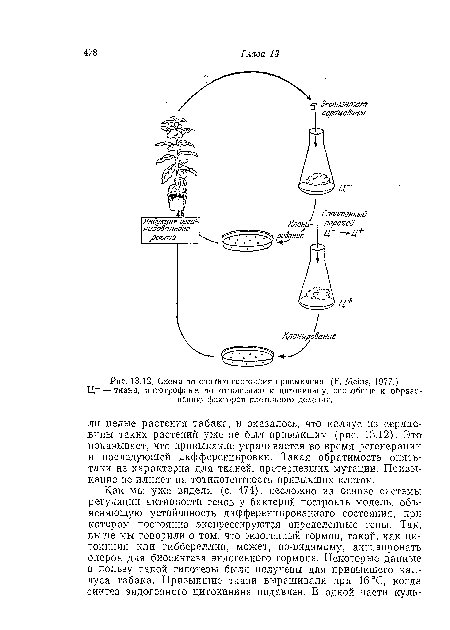 |
Далее
| Устойчивое положение перегородки с минимальной поверхностью мсладу двумя соприкасающимися равными пузырями. (D.A.W. Thompson, Growth and Form, Cambridge University Press, 1942.) |
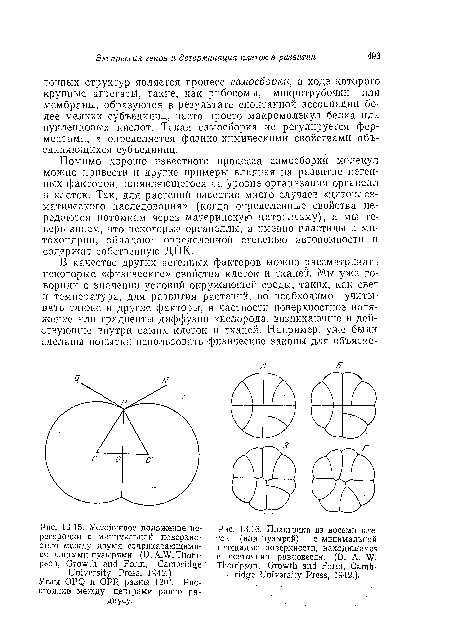 |
Далее
| Пластинка из восьми клеток (или пузырей) с минимальной площадью поверхности, находящаяся в состоянии равновесия. (D. A. W. Thompson, Growth and Form, Camb- |
|
Далее