Абсцизовая кислота может образовываться как прямым путем из мевалоновой кислоты, гак и при распаде виолаксантина — из промежуточного продукта ксанток-сина. При превращениях абсцизовой кислоты образуются фазеезая и дигидрофазеевая кислоты, а также конъюгаты, в том числе глюкозид. Молекулярный механизм действия состоит частью в изменении транспортных свойств мембран, частью в стимуляции или подавлении синтеза определенных ферментов или во влиянии на синтез РНК.[ ...]
Абсцизовая кислота была выделена из ряда покрытосеменных, голосеменных, папоротников и мхов, но ее, по-видимому, нет у печеночников. Вместе с тем у представителей по крайней мере восьми видов печеночников и некоторых водорослей была обнаружена лунуларовая кислота, которая у этих организмов, вероятно, играет физиологическую роль, аналогичную роли АБК у более высокоразвитых растений. Лунуларовая кислота является очень мощным ингибитором роста, и, кроме того, у некоторых печеночников она, возможно, участвует в регуляции роста почек и состояния покоя.[ ...]
Хотя абсцизовая кислота получила свое название за предполагаемую способность ускорять опадение (абсци-зию) листьев и плодов, в отделении листьев она, по-видимому, все же не играет никакой роли. Как уже говорилось, содержание абсцизовой кислоты в стареющих листьях или листовых черешках не находится в прямой связи с возрастанием тенденции к опадению. Предполагали, что по крайней мере в одном особом случае — при повышении уровня АБК и ускоренном опадении листьев в результате водного стресса у некоторых видов растений — должна существовать причинная связь между накоплением АБК й опадением. Однако более детальное изучение этого вопроса на хлопчатнике показало, что такой связи все же не существует. Более того, выяснилось, что водный стресс блокирует ток ауксина из пластинки к основанию листового черешка, так что главной причиной сбрасывания листьев в этом случае является, очевидно, недостаток ауксина в отделительной ткани.[ ...]
Кроме абсцизовой кислоты и родственного ей ксан-токсина встречаются и другие вещества, которые действуют как антагонисты ауксинов, гиббереллинов и цито-кининов. Их химическая структура может быть различной, а регуляторные функции с достоверностью не установлены.[ ...]
Обработка абсцизовой кислотой, наоборот, повышает тургор растений вследствие закрывания устьиц. Это влияние АБК подтверждает вывод о ее роли в движении устьиц, сделанный на основании изучения эндогенной АБК. Такие растения томатов содержат в десять раз меньше АБК, чем нормальные разновидности, и их устьица постоянно открыты. Однако если ежедневно опрыскивать завядающие мутанты томатов АБК, они полностью сохраняют тургор в течение нескольких дней и выглядят совершенно нормально. Помещение полосок эпидермиса с листьев удобных для таких опытов растений (в частности, СоттеИпа соттитэ) на растворы АБК также вполне определенно свидетельствует о том, что движение устьиц регулируется АБК путем подавления гормоном поступления К+ в замыкающие клетки и распада крахмала в хлоропластах этих клеток.[ ...]
Определение абсцизовой кислоты в растительном материале. Методы определения фитогормонов, ингибиторов роста, дефолиантов и гер бицидов».[ ...]
Идентификация абсцизовой кислоты. В работе Милборроу [3] приводится несколько требований, необходимых для идентификации этого ингибитора. Присутствие абсцизовой кислоты в растительном экстракте считается установленным, если: ингибитор присутствует в эфиррастворимой кислой фракции экстракта; значение этого ингибитора соответствует абсцизовой кислоты (метчика) в разных системах растворителей; признаки ингибирования (по биотесту) характерны для абсцизовой кислоты и отличаются от подавления, характерного для ингибиторов фенольной природы, таких, как салициловая кислота или кумарин; выделенный ингибитор обладает эффектом Коттона с характерной для абсцизовой кислоты кривой оптической вращательной дисперсии (ОРД).[ ...]
Таким образом, абсцизовая кислота может быть обнаружена не цветными реакциями, а с помощью биотестов и путем повторного хроматографирования элюатов в системах растворителей, для которых известно ее расположен 1е ( /).[ ...]
Таким образом, абсцизовая кислота может быть обнаружена не цветными реакциями, а с помощью биотестов и путем повторного хроматографирования элюатов в системах растворителей, для которых известно ее расположен 1е ( /).[ ...]
Регуляторная роль абсцизовой кислоты и ксантоксина в процессе роста в длину весьма сомнительна; это видж> из результатов сравнения карликовых и нормальных растений и изучения роста в длину, регулируемого светом.[ ...]
Данные о подвижности абсцизовой кислоты в нескольких смесях растворителей приведены в таблице.[ ...]
| Структурные формулы абсцизовой кислоты (слева) и ксантоксина (справа). |  |
Очень близкий по структуре к абсцизовой кислоте ксантоксин был открыт в 1968 г. Тейлором как сильный ингибитор прорастания. По ряду биологических свойств он в качественном и количественном отношении сходен с абсцизовой кислотой.[ ...]
| График доза—эффект для абсцизовой кислоты в тесте с отрезками колеоптилей овса и в тесте с ряской (ср. также рис.. 5.4). (По ТШЬе^.) | 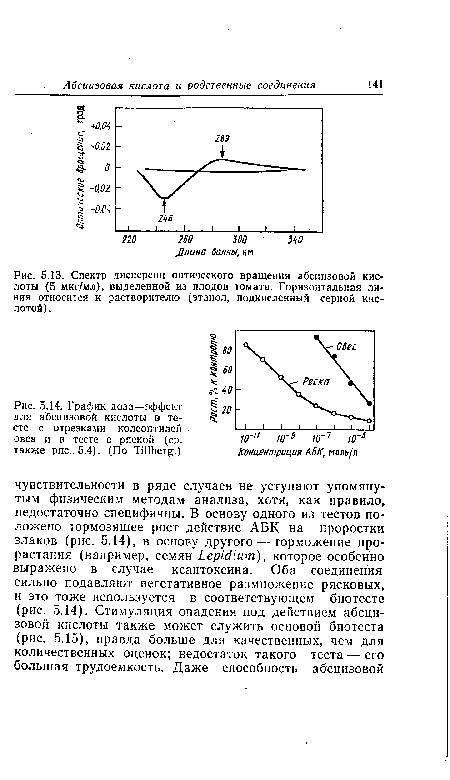 |
В противоположность цитокининам абсцизовая кислота и этилен у большей части растений ускоряют старение. Такой эффект можно наблюдать не только на изолированных листьях или листовых высечках, но и на листьях, сохранивших естественную связь с побегом. Под влиянием этилена ускоряется и созревание плодов, а также «отцветание», т. е. завядание и отмирание цветолистиков. Как в созревающих плодах, так и в стареющих органах цветка снижение содержания РНК и белков, повышение активности гидролитических ферментов и другие характерные для стареющих тканей процессы по времени связаны с повышением продукции этилена. Этилен, по-видимому, прежде всего повышает проницаемость мембран, 1то ведет к нарушению компартментализации и ускорению автолиза, причем одновременно усиливается синтез :амого этилена. Таким образом, процессы старения, которые вначале ограничены немногими клетками, могут Зыстро распространяться, в результате чего синхронизируется процесс старения во всем органе или же во всем астении.[ ...]
| Содержание ауксина, цитокинина и абсцизовой кислоты в ягодах винограда в сопоставлении с кривой их роста. (По А11е>теШ.) | 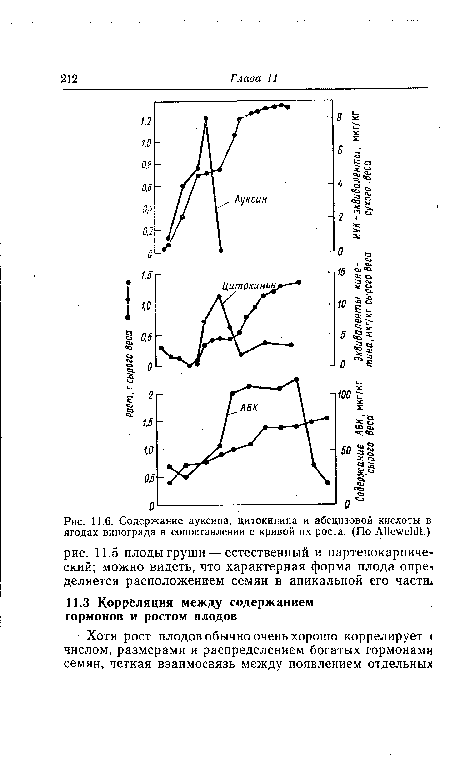 |
Располагая большими количествами абедизовой кислоты (порядка 1 мкг или более), можно воспользоваться высокоспецифическим методом определения, который основан на необычайно высокой оптической активности молекул АБК, связанной с асимметрией атома С-Г. Характеристический угол вращения составляет в сернокислом спиртовом растворе при 289 нм +24 000°, а при 246 нм —69 000°. Столь высокие значения лишь очень редко наблюдаются у других оптически активных веществ. Таким образом, спектр дисперсии оптического вращения (рис. 5.13) позволяет качественно и количественно определять абсцизовую кислоту.[ ...]
| Известные способы биохимической инактивации абсцизовой кислоты |  |
Многие эффекты проявляются только при взаимодействии абсцизовой кислоты с другими гормонами. Например, АБК тормозит растяжение клеток в отрезках колеоптилей овса только при наличии ауксина, индуцирующего рост клеток. Чаще всего АБК ведет себя по отношению к другим гормонам как антагонист, например по отношению к ауксинам при росте клеток растяжением, к цитокининам при клеточном делении (см. рис. 5.4), к гиббереллину при индукции синтеза амилазы. Механизм этих взаимодействий в большинстве случаев неизвестен. Возможно, что в антагонизме по отношению к ИУК при росте клеток растяжением существенную роль играет, как уже отмечалось, замедление выброса протонов. Однако маловероятно, чтобы АБК и ИУК. конкурировали ьа один и тот же рецептор; во всяком случае, нет точных фактов, указывающих на такого рода конкуренцию. Скорее АБК могла бы влиять на синтез или распад других гормонов. Судя по некоторым данным, распад гиббереллинов в алейроновом слое ячменя и синтез ауксина в колеоптилях, возможно, зависят от АБК.[ ...]
Однако после удаления листьев и декапитации содержание абсцизовой кислоты в расчете на одну почку не снижалось в той мере, как того следовало бы ожидать, если бы торможение было вызвано абсцизовой кислотой. Несмотря на это противоречие, гипотеза об участии абсцизовой кислоты в коррелятивном торможении почек весьма привлекательна. Мы уже упоминали, что покоящиеся почки многих растений отличаются повышенным содержанием абсцизовой кислоты (разд. 5.2.3.3). Коррелятивно заторможенные почки тоже можно рассматривать как покоящиеся (коррелятивный покой), и, как известно, у многих древесных растений коррелятивный покой может прямо переходить в состояние внутреннего зимнего покоя.[ ...]
Хотя в молодых листьях наряду с ауксином образуется много абсцизовой-кислоты и этилена, пока еще не ясно, влияют ли эти гормоны на рост листьев. Хотя абсци-зовая кислота тормозит рост изолированных листовых высечек, ее содержание в листе при нормальном водном режиме все же не коррелирует со скоростью роста. Однако не исключено, что появление очень маленьких листьев ксероморфного характера при сильном водном дефиците обусловлено резким повышением содержания АБК и одновременным снижением уровня цитокинина. В пользу такого рода связи свидетельствуют проведенные недавно эксперименты с листьями пшеницы. Подобное действие может, вероятно, оказывать и этилен.[ ...]
До сих пор нет окончательных данных о механизмах действия абсцизовой кислоты (АБК). Вне всякого сомнения этот ингибитор подавляет синтез белка, повышая активность PH Казн и снижая уровень РНК, но, как полагают, это не первичный механизм действия АБК. Другой возможный механизм — закрывание устьиц — быстрый мембранный эффект АБК, по-видимому, с синтезом РНК не связан. Весьма ценно, что, обсуждая взаимодействие отдельных гормонов и механизмы гистогенеза, регулируемые ауксинами, гиббереллинами и цитокинина-ми, авторы особое внимание уделяют корреляционным взаимодействиям органов.[ ...]
Казалось, что такое знание поможет понять механизм внутриклеточного действия ауксинов. Позднее, после открытия гиббереллинов, цнтокининов, этилена и абсцизовой кислоты, исследование этих гормонов шло теми же путями.[ ...]
Третьим регулятором в дополнение к ауксину и этилену служит абсцизовая кислота. Напомним, что она была открыта в связи с изучением ряда физиологических процессов, в том числе преждевременного опадения плодов у хлопчатника (разд. 5.2.1). У многих растений существует несколько фаз опадения плодов: 1) непосредственно после опыления, 2) к началу фазы роста (так называемое «июньское опадение» — в центральной Европе эта фаза у многих плодовых деревьев приходится на июнь и июль) и 3) во время созревания плодов. У хлопчатника опадение как незрелых, так и зрелых плодов связано с сильным повышением содержания абсцизовой кислоты (см. рис. 5.9). Это относится и к июньскому опадению плодов груши, а также к опадению молодых плодов у Lupinus и Phaseolus под влиянием более старых плодов. Эти данные позволяют думать, что более старые плоды влияют на содержание АБК в более молодых плодах. Неясно, однако, осуществляется ли такая коррелятивная зависимость просто путем передачи абсцизовой кислоты молодым плодам от старых или же каким-либо иным путем.[ ...]
Для простоты природное соединение (- -)-2-г с-АБК называют просто абсцизовой кислотой, или АБК.[ ...]
Динамика активности веществ со свойствами гиббереллинов, ауксинов и абсцизовой кислоты в листьях рудбекии изучалась в условиях 9-часового короткого дня, естественного длинного дня и в условиях непрерывной темноты [Кочанков и др., 1979].[ ...]
Кефели В.И., Коф Э.М., Власов П.В., Кислин Е.Н. природный ингибитор роста - абсцизовая кислота. М., Наука. 1989. 184 с.[ ...]
| 5.1 | 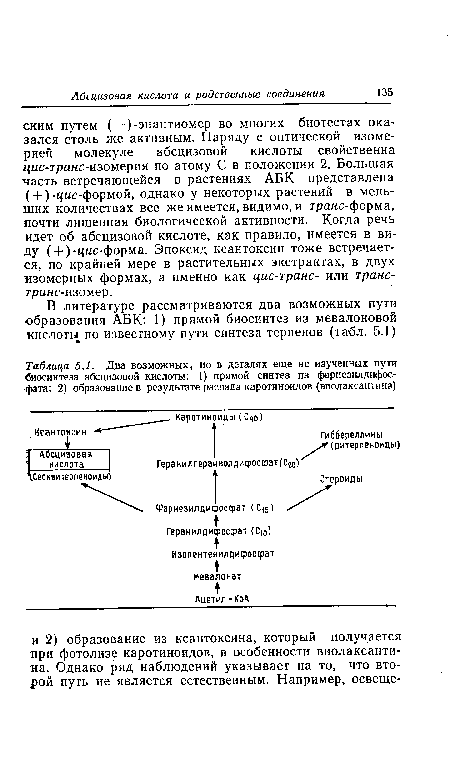 |
Дается краткий литературный обзор, касающийся выделения и определения абсцизовой кислоты в растениях. Приводится оригинальный метод выделения и идентификации абсцизовой кислоты из почек черной смородины.[ ...]
В отличие от уровней гиббереллинов, цитокининов и этилена концентрация абсцизовой кислоты, по-видимому, не зависит от фитохромной системы, однако эта система влияет на уровень ксантоксина. В проростках гороха, подвергнутых кратковременному воздействию красного или дальнего красного света, были обнаружены одинаковые количества абсцизовой кислоты, но ксантоксина было в первом случае в 2—3 раза больше. Сильное тормозящее действие красного света на рост побегов, вероятно, не связано с содержанием ксантоксина: это было продемонстрировано точными кинетическими исследованиями.[ ...]
Анализ определения активности гиббореллипов, цитокишшов и содержания абсцпзовой кислоты в разных участках стебля при прямом и обратном градиенте цветения показал, что кольцевание, смещая градиент цветения почек, в той же мере смещает градиент распределения гормонов по стеблю (рис. 140). И в том, и в другом случае наиболее активный рост и быстрое зацветание почек (при прямом градиенте в верхних и при обратном градиенте в нижних узлах стебля) происходят на фойе высокой активности гиббереллипа и цитокипина и низкого содержания абсцизовой кислоты.[ ...]
Этим методом можно обнаружить активность таких соединений, как индолил-3-уксусная кислота (ИУК), абсцизовая кислота, а также фенольные ингибиторы. Строение этих соединений приводим на стр. 8. Предлагаемый метод определения биологической активности свободных ауксинов и ингибиторов основан на хроматографическом разделении эфирного экстракта из растительного материала с последующей оценкой степени стимуляции и торможения отдельных зон хроматограмм при помощи биологических тестов. Ниже приводим описание реактивов последовательных процедур метода.[ ...]
| Высота проростков пшеницы в возрасте 14 дней после обработки гибберелловои кислотой и абсцизовой кислотой | 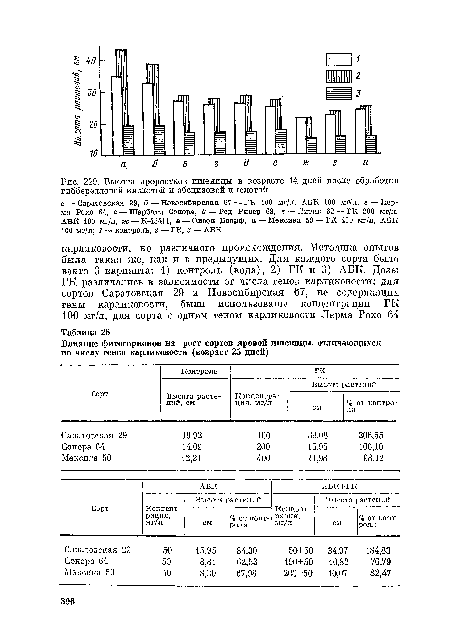 |
Сесквитерпеноидные соединения также содержат ингибиторы. Наиболее известна из них абсцизовая кислота (АБК). Покой семян яблони и почек явора (Acer pseu-doplatanus) связывается с присутствием АБК. Механизм действия таких веществ, как кумарин, заключается в блокировании или инактивации ферментов, необходимых для прорастания. Например, было установлено, что в присутствии ингибитора прорастания блокируется активность а и р-амилаз. Амилазы необходимы для гидролиза крахмала, т. е. превращения сложных нерастворимых углеводов в простые растворимые формы. Тормозящее действие абсцизовой кислоты приписывают подавлению ею активности веществ, стимулирующих прорастание, например гибберелловой кислоты. Аналогично покою, обусловленному непроницаемостью семенной оболочки, ингибиторы задерживают и удлиняют период прорастания. Вымывание дождевыми осадками и разложение микроорганизмами являются примерами того, как ингибиторы прорастания удаляются из плода и семенных оболочек.[ ...]
Из-за отсутствия метчика мы не могли сравнить величину Я/ выделенного ингибитора с Щ абсцизовой кислоты. Поэтому при сравнении этих показателей мы пользовались литературными данными. Несмотря на то, что значение величины Щ зависит от многих факторов, совпадение наших данных с литературными вполне удовлетворительно. Показатели/?/ абсцизовой кислоты и ингибитора из почек черной смородины в различных системах растворителей были следующими.[ ...]
В последнее время в тканях многих растений был обнаружен терпеноидный ингибитор роста — абсцизовая кислота, которая вместе с фенолами входит в состав ß-ингибитора. Абсцизовая кислота подавляет на 50% рост отрезков колеоптилей в дозе 0,08 мг/л, т. е. обладает в 100 раз более сильным действием на рост биотестов, чем фенольные ингибиторы. Хотя предлагаемый метод не предусматривает выделения отдельных чистых веществ, при выборе системы растворителей следует руководствоваться правилом выбирать такую смесь, которая позволила бы разделить природные регуляторы друг от друга на наибольшее расстояние. В таблице приводятся данные о разделении ИУК, гиббереллина, флоридзина, га-кума-ровой и абсцизовой кислот в нескольких смесях растворителей.[ ...]
Сравнение биологической активности ингибитора из почек черной смородины с биологической активностью абсцизовой кислоты, кумарина и салициловой кислоты показало, что ингибитор из почек черной смородины вызывает подавление роста, характерное для абсцизовой кислоты. Ингибитор из почек (1 г сухого вещества в пятне) и абсцизовая кислота в концентрации 10 4% вызывали полное подавление прорастания изолированных зародышей яблони в первые 2— 3 дня в термостате при 25°. После трех суток проращивания зародыши выносили на свет. На свету начиналось активное зеленение семядолей, образование придаточных корней, нормальное, но растянутое прорастание. Салициловая кислота и кумарин вызывали полное подавление прорастания изолированных зародышей яблони в концентрациях в 500 раз более сильных, чем абсцизовая кислота. При выставлении зародышей на свет нормального прорастания не было. В концентрациях, близких к гормональным, эти вещества ингибирующие действие не проявляли.[ ...]
Кроме гормонов-стимуляторов, как известно, существует обширный класс ингибиторов роста. Антагонистам фитогормонов, а именно абсцизовой кислоте (АБК) и этилену, посвящены специальные главы. АБК рассматривается как гормональный ингибитор роста, участвующий в регуляции покоя, опадения листьев и движений устьиц. Описаны два пути метаболизма АБК — превращение в сесквитерпены через фарнезилпирофос-фат и расщепление каротиноидных структур. В этой же главе специальный раздел посвящен и другим ингибиторам (в частности, фенольной природы), исследование которых проводится у нас в стране, о чем упоминает и автор.[ ...]
Кроме гормонов-стимуляторов, как известно, существует обширный класс ингибиторов роста. Антагонистам фитогормонов, а именно абсцизовой кислоте (АБК) и этилену, посвящены специальные главы. АБК рассматривается как гормональный ингибитор роста, участвующий в регуляции покоя, опадения листьев и движений устьиц. Описаны два пути метаболизма АБК — превращение в сесквитерпены через фарнезилпирофос-фат и расщепление каротиноидных структур. В этой же главе специальный раздел посвящен и другим ингибиторам (в частности, фенольной природы), исследование которых проводится у нас в стране, о чем упоминает и автор.[ ...]
Кроме гормонов-стимуляторов, как известно, существует обширный класс ингибиторов роста. Антагонистам фитогормонов, а именно абсцизовой кислоте (АБК) и этилену, посвящены специальные главы. АБК рассматривается как гормональный ингибитор роста, участвующий в регуляции покоя, опадения листьев и движений устьиц. Описаны два пути метаболизма АБК — превращение в сесквитерпены через фарнезилпирофос-фат и расщепление каротиноидных структур. В этой же главе специальный раздел посвящен и другим ингибиторам (в частности, фенольной природы), исследование которых проводится у нас в стране, о чем упоминает и автор.[ ...]
Рост и развитие растений подвержены регуляции со стороны фитогормонов (регуляторов роста растений), которые представляют собой сигнальные молекулы и которыми являются ауксины, гиббереллины, цитокинины, абсцизовая кислота и этилен. Перечисленные соединения либо синтезируются в клетках, либо транспортируются к клеткам-мишеням. Наибольший эффект этих соединений проявляется при их сочетанном действии. Например, ауксин, индолилуксусная кислота, стимулирует образование корней, но в сочетании с гиббереллином содействует росту корней в длину, а в сочетании с цитокинином — стимулирует закладку и рост боковых почек.[ ...]
Таким образом, картина апикального доминирования корня резко контрастирует с таковой для побега. В то время как в побеге ауксины из его верхушки тормозят рост почек, а цитокинины из корня стимулируют, в корне те же гормоны действуют как раз наоборот. Роль абсцизовой кислоты и родственных ей соединений в этих двух случаях апикального доминирования еще не ясна.[ ...]
Имеющиеся сведения о цветных реакциях на природные ауксины и ингибиторы говорят скорее о подсобном характере этого приема анализа, который может на первом этапе дать лишь общую характеристику расположения пятен на хроматограммах. Причем индольные ауксины и абсцизовая кислота на хроматограммах с помощью цветных реакций не обнаруживаются. Тем большее значение для обнаружения физиологически активных веществ приобретают биологические методы исследования (биотесты).[ ...]
Таким образом, выделенный нами из кислой фракции метаноль-ного экстракта почек черной смородины ингибитор по характеру биологической активности, Rf в различных системах растворителей, по характерным экстремумам кривой оптической вращательной дисперсии идентичен абсцизовой кислоте (АК).[ ...]
В растительном организме АБК образуется из мевалоповой кис-поты через изопентенилпирофосфат. Таким обравоы, путь образования АБК идет вначале через те же этаньт, что и образование фитогормона гиббереллина. Во многих случаях АБК и по своему действию является антагонистом гиббереллина. АБК содержится в различных органах растений, особенно в состоянии глубокого покоя. Она обнаружена в почках, сухих семенах, в клубнях картофеля. Баланс между фитогормонами л абсцизовой кислотой определяет способность данного органа к росту и морфогенезу. Имеются данные, что накопление АБК вызывает закрытие устьиц. Под влиянием АБК резко уменьшается проницаемость мембран, что, по-видимому, оказывает разностороннее тормозящее влияние на процессы обмена.[ ...]
На фазе повышенной устойчивости клетки наряду с накоплением протекторов перекисного нарушения мембран ускоряется синтез ряда низкомолекулярных защитных веществ. Хорошо известно, что под влиянием высокой и низкой температур, засоления, водного дефицита и т.п. в клетке многократно увеличивается количество имминокисло-ты пролина (Aspinale, Paleg, 1981). Ее ускоренный синтез, как и запуск биосинтеза антиоксидантов - пороговое явление. Индуктором накопления пролина, по-видимому, является абсцизовая кислота (Ке-фели и др., 1989). Молекула имминокислоты своей гидрофобной частью встраивается в мембрану, а торчащий наружу ее гидрофильный конец увеличивает водоудерживащее свойство мембран (Schobert, 1977). Последнее существенно для выживаемости клетки в условиях водного дефицита, причиной которого могут быть различные воздействия.[ ...]
Алейроновым слоем называют периферический слой клеток, содержащих большое количество белка и расположенных вокруг эндосперма семян злаков. Эта ткань очень активна при прорастании и на ранних стадиях роста проростков, а затем быстро дегенерирует и отмирает. До прорастания алейроновый слой служит запасающей тканью, а при прорастании он является источником ряда гидролитических ферментов, секретируе-мых в эндосперм и участвующих в мобилизации его запасных веществ. Таким образом, алейроновый слой представляет собой однородную ткань, состоящую из одинаковых клеток, запрограммированных на выполнение небольшого числа функций на ранних этапах жизни растения. После набухания семян, вышедших из состояния покоя, клетки алейронового слоя выполняют свои функции при условии получения ими соответствующих гормональных сигналов. Основным гормоном, регулирующим метаболизм алейронового слоя у семян ячменя, является гиб-береллин, поступающий из прорастающего зародыша. Одиако он, очевидно, вступает в сложное взаимодействие с абсцизовой кислотой и, возможно, с этиленом. Подавляющее большинство экспериментов было проведено на алейроновом слое семян ячменя, но такая же ситуация типична и для семян других злаков. Исключение составляют -семена пшеницы, у которых в регуляции начала гидролиза принимают участие также ауксины и цитокинины.[ ...]
Многие годы считалось, что затопление и засуха вызывают в растении ряд сходных физиологических изменений, и это легко понять, поскольку и в том, и в другом случае нарушается нормальное поступление воды в растения. В условиях засухи эти нарушения вызываются постепенным уменьшением влажности почвы, а при затоплении анаэробиоз в почве мешает нормальному функционированию корней, и их сопротивление водному току увеличивается. Однако недостаток воды в листьях при затоплении обычно не так велик, как во время засухи.. Вместе с тем засуха в отличие от затопления не вызывает эпи-иастии листьев и утолщения стебля. Эпииастия и утолщение стеблей затопленных растений возникают в результате повышенного содержания этилена в их побегах. Старение листьев у затопленных растений определяется как повышенным содержанием этилена, так и сниженным поступлением цитокининов из корней, страдающих от анаэробиоза почвы. Влияние этилена и цитокининов на старение листьев рассмотрено в гл. 12. Затопление корней также снижает поступление из корней в побег гнб-береллинов, и это служит причиной снижения в данных условиях роста стебля растяжением. Содержание абсцизовой кислоты в побегах затопленных растений слегка увеличивается, но не так сильно, как у растений в условиях засухи. Быть может, это связано с меньшим дефицитом воды у затопленных растений в сравнении с растениями, подвергнутыми воздействию засухи.[ ...]