Гаметангии — оогонии и антеридии — у диктиотовых тоже образуются на поверхности слоевища плотными группами (рис. 133, 2, 3). В оогониях развивается по одной яйцеклетке. Неоплодотворенные яйцеклетки способны прорастать партеногенетически, однако образующийся при зтом проросток вскоре погибает. Антеридии делятся горизонтальными и вертикальными перегородками на 150—1500 камер, в каждой из которых появляется по одному антерозоиду. Строение антерозоидов изучали у диктиоты дихотомической (Dictyota dicho-toma). Оказалось, что, в отличие от гамет и зооспор других бурых водорослей, они имеют по одному жгутику.[ ...]
Гаметангии многочисленные и разбросанные по поверхности гаметофита либо без всякого порядка, либо (у гроздовника) антеридии занимают вершину специального гребня, а ар-хегонии разбросаны по склонам гребня или на брюшной стороне гаметофита. Антеридии крупные, погруженные или слегка выступающие над поверхностью гаметофита; архегонии довольно глубоко погруженные.[ ...]
Гаметофит, гаметангии и оплодотворение.[ ...]
Очевидно, такие гаметангии не могли удовлетворить потребностям наземного существования. С переходом водорослевого предка высших растений к наземному образу жизни возникла прежде всего потребность в защите гамет от высыхания. Как в мужских, так и в женских гаметангиях самые наружные клетки должны были в зтих условиях стерилизоваться, т. е. потерять способность производить гаметы, и после соответствующих видоизменений превратиться в защитный слой. Предполагают, что именно таким путем, т. е. путем стерилизации наружного слоя клеток, развилась стенка ан-теридиев и архегониев.[ ...]
Покровпые листья окружают гаметангии и обычно отличаются от стеблевых размерами, формой, часто анатомическим строением, а иногда — окраской и консистенцией.[ ...]
| Защитные образования гаметангиев и развивающихся спорогонов у юнгерманниевых. Различные типы периантиев и их схематические разрезы | 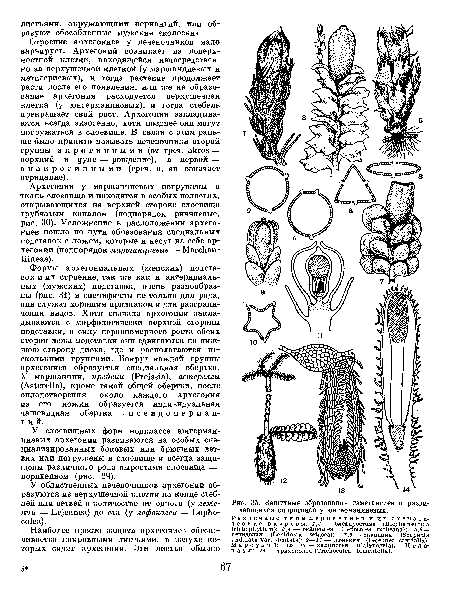 |
Органы полового размножения (гаметангии), как женские (архегонии), так и мужские (антеридии), чаще располагаются не поодиночке, а группами, нередко с многочисленными стерильными нитями (п а р а ф и з а м и). Обычно скопления гаметангиев окружены специальными, иногда сильно видоизмененными листовидными выростами или другими защитными образованиями. Гаметангии могут находиться на специальных подставках из видоизмененных побегов или их частей, а также могут быть погруженными в ткань слоевища.[ ...]
Органы полового размножения (гаметангии), как женские (архегонии), так и мужские (антеридии), чаще располагаются не поодиночке, а группами, нередко с многочисленными стерильными нитями (п а р а ф и з а м и). Обычно скопления гаметангиев окружены специальными, иногда сильно видоизмененными листовидными выростами или другими защитными образованиями. Гаметангии могут находиться на специальных подставках из видоизмененных побегов или их частей, а также могут быть погруженными в ткань слоевища.[ ...]
| Прорастание споры, строение заростков (гаме-тофитов), гаметангиев и зародыша у хвоща (ЕдшзеЬит) |  |
С усилением половой дифференциации количество мужских гамет в гаметангии возрастало. Так получился антеридий высших растений с его многочисленными мелкими сперматозоидами. Количество женских гамет, напротив, уменьшилось. Таким путем возник в конце концов архегоний с одной большой и неподвижной яйцеклеткой. В эволюции архегония стерилизация сказалась весьма сильно, а сама женская гамета лишилась жгутиков и стала неподвижной. Шейковая и брюшная канальцевые клетки архегония также были когда-то гаметами, но в процессе эволюции стерилизовались. У разноспоровых растений в связи с редукцией их гаметофитов редуцируются и видоизменяются также аптеридии и архегонии, а у голосеменных родов вельвичия и гнетум и у всех цветковых растений они уже вовсе не образуются.[ ...]
Гаметофит строматоптериса подземный, бесцветный, длиной около 2 см. Гаметангии и ризоиды на его поверхности распределены равномерно.[ ...]
Гаметофиты гельминтостахиса короткие, в нижней части лопастные, в верхней, несущей гаметангии — цилиндрические.[ ...]
У антоцеротовых органы полового размножения развиваются эндогенно в ткани слоевища. Гаметангии неравномерно разбросаны в средней части слоевища; часто женские и мужские оказываются на одном и том же растении.[ ...]
Гаметангии представляют отростки . клеток мицелия, содержащие по одному ядру, они сливаются, зигота отделяется перегородками от оснований гаметангиев и превращается в восьмиспоровую сумку.[ ...]
Слоевище водорослей этого порядка лишено перегородок, только в основании ветвей и изредка гаметангиев возникают своеобразные пробки. В пристенном слое цитоплазмы содержатся многочисленные ядра и хлоропласты с пиреноидами или без них.[ ...]
Морфологически дифференцированных гаметангиев нет. Затем образуется маленький клубок гиф, в дальнейшем дифференцирующийся на шляпку и ножку. Зачаток апотеция в этот момент не превышает 1 мм.[ ...]
У низших аскомицетов (подкласс Негтавсо-mycetidae) половой процесс сходен с зигогамией у зигомицетов. Гаметангии разного пола морфологически сходны или малоразличимы и представляют выросты или веточки мицелия. После их слияния сразу происходит кариогамия и сумка развивается непосредственно из зиготы. Однако, в отличие от зигомицетов, в многоядерных гаметангиях сливаются только два ядра (нет множественной кариогамии), зигота не переходит в состояние покоя, а сразу развивается в сумку. В цикле развития низших аскомицетов, следовательно, есть только гаплоидная и диплоидная стадии (рис. 52).[ ...]
Для высших аскомицетов (подклассы Еиавсо-тусеШае и Ьоси1оа8сотусеШае) характерны дифференциация и усложнение строения гаметангиев. Образуются одноклеточный антеридий и аскогон, обычно с трихогиной. При оплодотворении содержимое антеридия по трихогине переходит в аскогон. После плазмогамии гаплоидные ядра разного пола не сливаются сразу, а объединяются попарно, образуя дикарионы. Из аско-гона вырастают аскогенные гифы, в которых ядра дикариона синхронно делятся. На концах аскогенных гиф развиваются сумки (см. рис. 50). Конечная клетка аскоген-ной гифы загибается крючком, ядра дикариона располагаются в месте перегиба и одновременно делятся. Пара ядер разного пола остается в месте перегиба крючка, одно ядро переходит в его кончик, а другое — в основание. Затем образуются две перегородки, отделяющие одноядерные конечную и базальную клетки крючка. В результате слияния этих клеток восстанавливается дикарион и может происходить повторное образование крючка. Средняя двухъядерная клетка крючка развивается в сумку. Она увеличивается в размерах, ядрадикариона сливаются. Образовавшееся диплоидное ядро делится редук-ционно, за мейозом следует еще одно, митотическое деление, и вокруг восьми гаплоидных ядер формируются аскоспоры.[ ...]
Половой процесс — анизогамия, как и у других сифоновых. Гаметы образуются в конечных веточках, которые, перед тем как стать гаметангиями, не меняя формы, отделяются перегородкой от остального слоевища. Перегородки образуются как внутренние кольцеобраз-пые утолщения боковой стенки в основании веточек. Эти утолщения, разбухая, оставляют в середипе лишь узкий тяж протоплазмы, но постепенно и этот тяж прерывается, так как цитоплазма веточки и главной оси одевается собственной оболочкой, а первоначальное утолщение остается как студневидная пробка между этими оболочками. Выход гамет осуществляется в результате ослизнения и растворения оболочки на вершине веточки или через боковое отверстие в боковой стенке.[ ...]
Происхождение наземных растений иногда пытаются связывать с бурыми водорослями из-за существования у них многогнездных спорангиев и гаметангиев, которые ошибочно принимают за многоклеточные органы размножения. Вместилища бурых водорослей делятся на камеры в последний момент при формировании в них зооидов, чего не наблюдается у наземных растений. В то же время бурые водоросли сильно отличаются от наземных растений набором пигментов, запасными питательными веществами, химическим составом оболочки.[ ...]
У современных представителей порядка известно только половое размножение. Половой процесс — изогамия. Гаметы образуются в специальных гаметангиях. У большинства дазикладовых гаметангии возникают как шаровидные выросты на вершине или сбоку ветвей первого порядка и соответствуют видоизмененным ветвям второго порядка (рис. 232, 3, 4). У аце-табулярии гаметангии образуются на коротких сегментах первого порядка как специальные выросты (рис. 232, 9). Внутри гаметангия образуются толстостенные многоядерные цисты со специальной крышечкой (рис. 232, 10). Зрелые цисты содержат множество гамет. При разрушении стенок гаметангия цисты выходят во внешнюю среду и уже здесь из них высвобождаются гаметы. Цисты, а не зиготы могут служить покоящимися стадиями. Существует мнение, что образование цист связано с обызвествлением слоевища. У некоторых лишенных извести родов их нет и гаметы образуются непосредственно в гаметангиях.[ ...]
Наряду с этим существует другой вопрос. Но здесь они образуются иными способами: а) вследствие впячивания поверхностного слоя клеток слоевища за счет активного деления и роста клеток в отдельных точках его поверхности; б) в результате развития органов размножения на каком-то участке и обрастания его окружающими вегатативными тканями; в) путем появления органов размножения на клетках слоевища, выстилающих полость, которая перед этим образовалась вследствие разрушения клеток (некоторые известковые красные водоросли).[ ...]
Основными элементами в этом способе размножения являются копулирующие отроги, выполняющие половую функцию, и зигоспора — продукт полового процесса.[ ...]
Пельвеция желобчатая (P. canaliculata) имеет слоевище в виде кустов высотой до 15 см с дугообразными ветвями, согнутыми по продольной оси в виде желоба. Гаметангии созревают с конца июня по сентябрь. Растет пельвеция желобчатая пад поясом фукусов в среднем и верхнем горизонтах литорали. Распространена она от Белого и Баренцева морей до севера Португалии.[ ...]
В отличие от других многоклеточных водорослей у бурых водорослей, наряду с обычными одногнездными спорангиями (рис. 121, 2), имеются многогнездные спорангии и гаметангии, неправильно называемые многоклеточными (рис. 128, 1 а). Перед образованием зооспор или гамет содержимое многогнездных вместилищ делится тонкими перегородками на камеры, в которых оказываются заключенными по одному ядру с участком цитоплазмы. В каждой камере развивается по одной, реже по две зооспоры или гаметы. На поверхности слоевища многих бурых водорослей развиваются особые многоклеточные волоски, имеющие вид нити из одного ряда клеток с зоной роста при основании; клетки зоны роста делятся чаще других и поэтому имеют мелкие размеры (рис. 121, 1 б).[ ...]
Основными функциями листьев являются фотосинтез и удержание влаги. Специализированные покровные листья вместе с п е р и а н-т и е м (от греч. peri — возле и anthos — цветок) защищают гаметангии и молодые спорого-ны от иссушения.[ ...]
Основные отличительные особенности циклоспоровых связаны с их циклом развития и размножения. Это, во-первых, отсутствие двух самостоятельно растущих форм развития. Во-вторых, гаметангии развиваются в углублениях на слое клеток, который возникает из одной клетки, именуемой инициальной клеткой кон-цептакула, но мы ее будем называть проспорой. В-третьих, у всех до сих пор изученных циклоспоровых мейоз происходит при образовании гамет.[ ...]
Ассимиляционная ткань из вертикальных столбиков, клеточных тяжей или воздушных камер с устьицами. В коробочке, кроме спор, иногда имеются питательные клетки.[ ...]
Таким образом, развитие спор полового размножения у диподасковых имеет большое сходство с наблюдаемым у зигомицетов. Как и у зи-гомицетов, здесь может происходить слияние многоядерных гаметангиев (Dipodascus albidus), сменяющихся в процессе эволюции одноядерными (Dipodascus uninucleatus). Однако при слиянии многоядёрных гаметангиев диподасковых не происходит множественной кариогамии (слияние ядер), а сливаются только два ядра. Здесь следует отметить, что уже у некоторых зигомицетов (род Endogone) при гаметангиога-мии сливаются только два ядра, а остальные ядра многоядерных гаметангиев дегенерируют. Сумка аскомицетов гомологична зародышевому спорангию зигомицетов, у диподасковых же их сходство увеличивается еще и тем, что количество аскоспор в сумке может достигать нескольких десятков и не фиксировано строго, как у остальных аскомицетов.[ ...]
У метцгериевых (Metzgeriaceae) слоевище мелкое, светло-зеленое, дихотомически или мо-ноподиально разветвленное, узкое, с однослойной пластинкой и многослойной жилкой. Поверхность слоевища с волосками. Гаметангии — на специальных веточках с нижней стороны слоевища. Колпачок мясистый, покрыт волосками. В семействе один род метцгерия (Metzge-па, рис. 29), в котором насчитывают свыше 120 видов, распространенных преимущественно в тропиках. В СССР — 4—5 видов.[ ...]
Мужские и женские органы полового размножения (антеридии и архегонии) у мхов чаще всего собраны в группы, обычно окруженные специализированными, иногда окрашенными покровными листьями. Возникают гаметангии или на верхушках главных побегов, или на очень коротких боковых ветвях. Между анте-ридиями и архегониями часто находятся нитевидные или булавовидные парафизы.[ ...]
Таким образом, у бриопсиса мы видим уже специализированные органы полового размножения, но, поскольку они сохраняют ту же морфолох ию, что и вегетативные веточки, молено считать, что дифференциация гаметангиев здесь еще не закончена.[ ...]
Цикл развития кутлериевых заключается в чередовании диплоидных спорофитов и гаплоидных гаметофитов, которые бывают одинакового или разного строения. Половой процесс у кутлериевых — гетерогамия. Мно-гогнездные гаметангии развиваются на разветвленных или неразветвленных однорядных выростах (рис. 130, 2, 3). Женские гаметангии с более крупными камерами, чем мужские. Женские гаметы соответственно крупнее мужских, содержат несколько хлоропластов и двигаются менее продолжительное время. Гаметы у кутлериевых высвобождаются через отверстие в каждой камере. Слияние их происходит после того, как женская гамета прикрепилась к грунту или другим водорослям.[ ...]
Образование зооспор и гамет у бурых водорослей происходит во вместилищах двух основных типов: одногнездных (рис. 121, 2) и много-гнездных (рис. 128, 1 а). Многогнездные вместилища могут функционировать как спорангии и как гаметангии. Внешне они при этом не отличаются так же, как зооспоры и гаметы. Одногнездные вместилища чаще бывают спорангиями. Мейоз у бурых водорослей происходит при образовании спор в одногнездных спорангиях, лишь у циклоспоровых он приходится на момент образования гамет.[ ...]
Наиболее широко распространена диктиота дихотомическая. Ее слоевища образуют большие дерновинки высотой до 20 см, ширина ветвей достигает 4—8 мм. Диктиота дихотомическая интересна четкими периодами в образовании и созревании гаметангиев. У берегов Англии гаметангии начинают развиваться в период квадратурных приливов, а окончательное созревание и выход гамет происходит в течение нескольких приливов, следующих после самого высокого сизигийного прилива. При этом гаметы созревают каждые две недели. Такая периодичность в размножении и связь его с лунным ритмом сохранялась при выдерживании слоевищ в лаборатории в течение нескольких месяцев. У Атлантического побережья Северной Америки созревание гамет происходит с интервалом в один месяц. Развитие гамет начинается за день до сизигийного прилива в полнолуние или на следующий день, гаметы созревают 6 или 8 дней спустя. В заливе Петра Великого (Японское море) обычно чаще встречаются растения с тетраспорангиями, которые иногда все превращаются в проростки па материнских растениях, и ветви последних становятся лохматыми, так как проростки, прежде чем отделиться, вырастают высотой в несколько миллиметров. Диктиота дихотомическая растет в верхней сублиторали в местах с движением воды.[ ...]
Род кутлерия (СиПепа) пользуется наибольшей известностью (рис. 129, 3, 4). Он служит классическим примером гетероморфной смены форм развития. У гаметофита слоевище однолетнее, вертикальное, вееровидное или кустистое, высотой до Ъсм. Гаметангии развиваются на разветвленных или простых однорядных выростах. Слоевище спорофита однолетнее или многолетнее, корковидное, диаметром до 10 см. В спорангиях образуется от 8 до 32 зооспор. Раньше, до того как был изучен цикл развития кутлерии, ее спорофит считали особым родом.[ ...]
Женские гаметы содержат многочисленные хлоропласты с пиреноидами; мужские — всего 1—3 хлоропласта, лишенные пиреноидов. Половой процесс, как правило, анизогамия. Дифференцированные гаметангии образуются как боковые выросты грушевидной формы на пузырях и отделяются от них специальными слизистыми перегородками, как у бриопсиса. Когда гаметы созревают, оболочка на вершине гаметангия разрывается, и гаметы выходят в окружающую среду. Зигота прорастает обычно без периода покоя. Новое поколение опять дает гаметы, и цикл развития завершается.[ ...]
Фукус зубчатый (F. serratus) выделяется среди остальных видов ветвями с зубчатыми краями. Слоевища его раздельнополые, в длину достигают м. Ъ условиях Баренцева моря гаметы созревают в августе — сентябре, единичные рецептакулы со зрелыми гаметангия-ми встречаются на отдельных растениях почти круглый год, крайне редко их можно найти весной. Фукус зубчатый растет в нижнем и иногда в среднем горизонте литорали в верхней сублиторали. Встречается у берегов Европы от Новой Земли до севера Португалии и у берегов Северной Америки в районе залива Святого Лаврентия.[ ...]
Род псеедобриопсис (Pseudobryopsis) обладает наиболее высокой организацией в семействе. По строению и внешнему виду он очень сходен с бриопсисом, но отличается наличием перегородок в основании вегетативных веточек и более дифференцированными гаметангиями. Веточки, в которых образуются гамоты, видоизменяются, приобретая урповидную или грушевидную форму. Освобождаются гаметы через верхушечную пору (рис. 230, 11).[ ...]
Бесполое размножение осуществляется при помощи зооспор с одним волочащимся сзади жгутиком. Половое размножепие представляет собой слияние подвижных гамет или одинаковых по величине и строению (изогамия), или различных (гетерогамия), образующихся в гаметангиях. При этом подвижные клетки (зооспоры, гаметы) бластокладиевых отличаются от зооспор и гамет других грибов тем, что они более активно движутся, а также наличием хорошо заметной ядерной шапочки, состоящей из массы РНК (см. рис. 4).[ ...]
У всех циклоспоровых, кроме аскозейры, наблюдается оогамия, В антеридиях образуется по 64 антерозоида, в оогониях развивается 1—8 яйцеклеток. Мейоз происходит при первых двух делениях ядра. В оогониях, несущих даже одну яйцеклетку, всегда образуется 8 ядер. Гаметангии циклоспоровых выглядят одногнездными, однако у некоторых представителей в антеридиях на стадии 32 ядер наблюдали нежные перегородки. В оогониях некоторых циклоспоровых яйцеклетки разделены нежными перегородками (хормозира, кси-фофора) или грубыми (пельвеция желобчатая). Эти факты свидетельствуют о том, что гаметангии циклоспоровых, видимо, все-таки про изошли от многогнездных вместилищ других бурых водорослей.[ ...]
Содержимое споры представляет собой первую клетку гаметофазы, и из нее в результате ряда митотических делений развивается гаме-тофит. В отличие от обоеполых гаметофитов однополые гаме-тофиты, особенно мужские, состоят из гораздо меньшего числа клеток и характеризуются возрастающей в процессе эволюции высших растений редукцией гаметангиев.[ ...]
У двух других видов — одноядерного дипода-скуса (Dipodascus uninucleatus), обнаруженного в Канаде и США на куколках фруктовой дрозофилы, и собранного диподаскуса (Dipodascus aggregatus), встречающегося в Европе и Северной Америке в личиночных галереях короедов, — мицелий состоит из одноядерных клеток. Гаметангии также одноядерны.[ ...]
У верхоплодных мхов стебель прямой или почти прямой и обычно слабоветвистый. У большинства видов ветви развиты только в верхней части стебля. Архегонии, а позднее и спорогоны возникают у них на верхушке стебля или основных ветвей; на развитие га-метангиев обычно тратится верхушечная клетка , и поэтому апикальный рост стебля и ветвей прекращается. У этих мхов, как правило, новые боковые ветви (инновации) образуются под гаметангиями и растут в направлении главного стебля, а на верхушке могут снова давать гаметангии. При симподиальном типе ветвления эти побеги возникают по одному; при вильчатом ветвлении или ложной дихотомии образуются парные побеги; у некоторых мхов возникает по нескольку побегов, т. е. имеет место пучковидное ветвление (например, у сфагнума, табл. 6).[ ...]
Цистозейра толстоногая (С. сга8з1рез), произрастающая в дальневосточных морях СССР, имеет слоевища иного облика, чем предыдущие виды. Ствол у нее короткий и несет на верхнем конце короткие и толстые, иногда булавовидные, многолетние ветви, на которых вырастают длинные ветви неограниченного роста с короткими боковыми веточками. Слоевище в длину достигает 2—6 м. В нижней части слоевища бывают узкие «листья» без ребер. Воздушные пузыри одиночные или цепочками располагаются на конечных ветвях. Гаметангии созревают на юге ареала в июне, на севере — в августе.[ ...]
У одних представителей этого семейства наблюдается типичный для высших аскомицетов половой процесс (например, у Мопавсив ригригеив). Однако у многих эуроциевых происходит его морфологическая редукция. В этих случаях в дикарионы объединяются ядра самого аскогона.[ ...]
Род аскофиллум (Ascophyllum) включает один вид аскофиллум узловатый (A. nodosum). Слоевище его, достигающее 1—1,5 м в длину, имеет длинные редкодихотомически-ветвящиеся ветви, на которых попеременно или супротивно расположены более короткие ветви. Рецептакулы развиваются как короткие булавовидные веточки по бокам крупных; при созревании гамет они раздуваются на вершине и становятся похожими на крупные ягоды. Мужские экземпляры более темной окраски, с менее округлыми ветвями и более мелкими рецептакулами. Веточки-рецептаку-лы появляются на слоевищах с середины июля до конца октября, тогда же в них развиваются проспоры и гаметофиты. Первые гаметангии возникают в конце декабря, гаметы созревают в июне, и в течение месяца происходит их выход. Аскофиллум растет в нижнем и отчасти в среднем горизонте литорали и в верхней сублиторали (в Белом море). Распространен аскофиллум у берегов Азии и Европы от Карского моря до Португалии и у берегов Америки от Баффиновой Земли до штата Нью-Джерси (США).[ ...]
Одна из наиболее примитивных форм зеленого гаметофита характерна для семейства ма-раттиевых. Гаметофиты мараттиевых мясистые, довольно долговечные (иногда живут по нескольку лет), достигают довольно значительных размеров (иногда длиной более 2—3 см). Более старые гаметофиты могут на верхушке дихотомически ветвиться, что, несомненно, является примитивным признаком. Гаметофиты мараттиевых бывают как продолговатыми, так и сердцевидными, причем первый тип может рассматриваться как более примитивный. Интересно, что антеридии на гаметофитах мараттиевых иногда могут развиваться также на верхней стороне, что, как предполагают некоторые авторы, вероятно, свидетельствует о происхождении этих гаметофитов из цилиндрических, у которых гаметангии рассеяны по всей поверхности.[ ...]
Эта теория благодаря авторитету Фритча нашла многих сторонников, к тому же до недавнего времени лишь она учитывала факт развития выстилающего слоя каждого концептакула из одной клетки. В довершение всего было обнаружено, что у аскозейры развитие концептакула и его выстилающего слоя начинается с деления одной клетки, лежащей на поверхности слоевища, при этом образуется узкая трубка, которая подобно паразитическому организму врастает внутрь, проходит через плотную кору и промежуточный слой и дорастает до сердцевины. Здесь среди рыхлорасположенных клеточных нитей конец трубки разрастается в полый шар — концептакул. То, что гаметофит у циклоспоровых получается диплоидным, не находится в противоречии с явлениями, наблюдаемыми у других водорослей.[ ...]
Класс улотриксовых объединяет очень большое количество крайне разнообразных зеленых водорослей, общим признаком которых является нитчатое или пластинчатое строение слоевища, состоящего у подавляющего большинства видов из нескольких или многих одноядерных клеток (исключение — многоядерная сфе-ротглея). Однако форма слоевища здесь может быть очень разнообразной. В простейшем случае это нить, образованная одним рядом клеток. Именно такая форма служит основой и начальной стадией всех последующих более сложных форм. Нити у одних видов свободно-живущие; у других — прикрепленные, и тогда дифференцированные на основание и вершину. Они могут быть ветвящимися и неветвящимися, равномерно цилиндрическими и сужающимися к вершине, распростертыми по субстрату и прямостоящими, иногда и теми и другими в пределах одного слоевища (разнонитчатое, или гетеротрихальное, строение), одиночно растущими, образующими скопления или дерновины и соединенными в слизистые колонии. Пластинчатые формы бывают однослойными и двухслойными или местами многослойными, распростертыми по субстрату и прикрепленными только в одном месте, плоскопластинчатыми и мешковидными. Размеры их также разнообразны — от микроскопических нитей, шириной в несколько микрометров до пластинчатых макрофитов с площадью пластин в сотни квадратных сантиметров. В их клетках находится по одному, реже по нескольку хлоропластов. Хлоропласты различной формы, преимущественно пластинчатые, пристенные, реже осевые. Половое размножение улотриксовых в основе однотипно, но строение гаметангиев весьма разнообразно.[ ...]