Пелагические икра и личинки барабули— Mutlus barba-tus L.— в Черном море выносятся течениями от берегов в открытое море, где бчльше пищи и меньше врагов, и только по достижении стадии, на которой они переходят к питанию бентосом, эти рыбы возвращаются в прибрежную зону.[ ...]
Пелагические эхоотметки на полигоне составляли от 68,2 до 97,3 %, т. е. подавляющая масса рыб во все времена года находилась здесь в движении, совершая перемещения вверх или скатываясь вниз по течению. Местные донные рыбы, дающие характерные отметки на эхограммах, распределялись в основном в зонах литорали и на склонах подводных ям и возвышенностей, где в результате сбоя потоков воды скорости течения ниже. Особой по условиям обитания оказалась зона круговоротов, здесь обнаружено большое число эхоотметок в толще воды и у дна. Эти зоны являются местами отдыха мигрирующих рыб и служат постоянным местообитанием ряда немигрирующих Видов как донных, так и пелагических рыб.[ ...]
У пелагических рыб, например кильки, наблюдается смешивание группировок и в вертикальном направлении. Этого рода смешивание рассматривается в той же работе А. К. Токарева.[ ...]
Для пелагических рыб, таких как лососи, острое зрение является важным условием выживания. Считается также, что диаметр глаза связан с максимальной высотой тела рыбы (Алеев, 1963), которая в свою очередь зависит от длины тела.[ ...]
| Пелагические икринки рыб (масштаб не соблюден) | 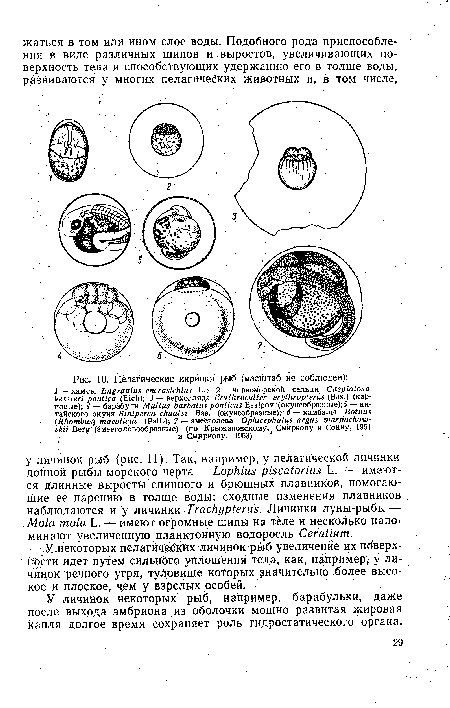 |
Хищную рыбу условно делят на донную — осетровые, сомовые, налим; пелагическую — лососевые, судак; зарос-левую — щука. Занимая разные экологические ниши, хищники при определенной плотности не конкурируют в питании. Для донных хищников кормом служат бычок, пескарь, гольян, шиповка и другие придонные мелкие формы; для пелагических — уклея, тюлька, килька, атерина; для за-рослевых — красноперка, горчак, плотва. Продуктивность хищных рыб колеблется от 0,2 до 2 ц/га.[ ...]
Многие пелагические личинки морских рыб совершают суточные вертикальные кормовые миграции, следуя в своих перемещениях за вертикальными перемещениями своих кормовых объектов — планктонных беспозвоночных, которые днем уходят на большие глубины, а ночью поднимаются к поверхности.[ ...]
Морские рыбы. Включают около 11,6 тыс. ёидов, живущих в морской воде. Среди них пелагические рыбы, живут в толще воды (пеламида, анчоусы, скумбрия, тунцы, луна-рыба и др.), и донные, которые живут у дна (камбалы, скаты, подкаменщики и др.).[ ...]
| Пелагическая рыба из семейства скумбриевых из верхних слоев миоценовых диатомитов. Остатки ископаемых рыб встречаются часто в богатых органическим веществом осадках. Монте-Каподарсо, центральная Сицилия. Длина масштабной полоски 2 см. |  |
Плотность пелагических рыб (горизонты от 3 до 8 м) в районе полигона также была выше. Основу этих рыб составляли тюлька и разновидовая молодь, размером менее 5—7 см.[ ...]
У некоторых пелагических личинок рыб увеличение их поверхности идет путем сильного уплощения тела, как, например, у личинок речного угря, туловище которых значительно более высокое и плоское, чем у взрслых особей.[ ...]
Плодовитость рыб намного больше, чем наземных позвоночных животных. Однако она сильно варьирует в зависимости от вида рыб. Наиболее плодовиты рыбы, откладывающие плавающую пелагическую икру. Меньшую плодовитость имеют рыбы, откладывающие икру на растениях. У рыб, охраняющих или прячущих свою икру, плодовитость невелика. У одного и того же вида плодовитость может сильно изменяться в зависимости от размера и возраста рыб, а также от условий питания.[ ...]
Сопротивление рыб сносу течением связывается с их двигательной активностью и может быть охарактеризовано тремя основными показателями: пороговыми скоростями течения, плавательной способностью и критическими скоростями течения, в пределах которого Возможна реореакция рыб. По результатам лабораторных исследований поведения пелагических рыб в потоке воды у большинства рыб выявлена четкая зависимость их распределения от гидравлической структуры потока (Павлов, 1979). При неустойчивом направлении течения, образовании вихревых зон одни рыбы уходят в сторону меньших скоростей течения, к области круговорота, другие имеют явную реореакцию и активно продвигаются против течения.[ ...]
В свою очередь, рыбы вод текучих (реофильные) и стоячих (лимнофильные) подразделяются на пелагических — обитателей толщи воды, придонных и донных.[ ...]
Если у взрослых рыб уменьшёНйе удельного веса обеспечивается, главным образом, за счет плавательного пузыря, то у икринок и личинок рыб это достигается иными способами (рис. 10).[ ...]
Питаются мелкими пелагическими рыбами и их молодью, ракообразными, крылоногими моллюсками. Сведения о биологии тихоокеанских лососей приведены в таблице 47.[ ...]
Размерный состав рыб на полигонах хорошо подтверждается машинным анализом эхозаписей. Так, осенью основу пелагических скоплений здесь составляют разновидовые особи размером 30— 50 мм (табл. 23).[ ...]
Сетное орудие лова рыбы. Различают неводы закидные (в морском, речном и озерном рыболовстве), закидываемые с берега и затем вытягиваемые на него, и обкидные (в море). Последние делятся на пелагические (лов в толще воды) и донные (выбрасываются с судна в воду и поднимаются на его борт). Разновидности пелагических неводов: кошельковые, распорные и кольцевые.[ ...]
В модели сообщества пелагических рыб оз. Дальнего (разделы 7.2 и 7.3) осуществлено практическое приложение многих рассмотренных в настоящей работе теоретических разработок к достаточно сложному, но хорошо изученному природному объекту.[ ...]
Амур — теплолюбивая рыба —больше распространен на юге. Половозрелым становится в возрасте 6-7 лет, в Краснодарском крае — в 3-5 лет. Плодовитость самок составляет 100-800 тыс. икринок, в среднем 500 тыс. икринок. Амур нерестится при температуре 26—30 °С. Икра пелагическая. Развитие происходит в толще воды. Амур в прудах созревает, но не нерестится. Поэтому его размножают заводским способом.[ ...]
Экологическая группа рыб, живущая (обитающая) в толще воды или на ее поверхности. Среди пелагических организмов различают пассивно плавающих на поверхности воды (плейстон) или в толще вод (планктон) и активно плавающих (нектон). Для пелагических организмов характерны прйспособления, обеспечивающие им плавучесть, например, плавательныё пузырь у рыб.[ ...]
Однако общая биомасса пелагических рыб в районе незначительно отличалась от прежних лет. В 1987—1988 гг. произошла вспышка численности синца от высокоурожайных поколений. Скопления этого вида дали существенную прибавку к общей биомассе пелагических рыб, в значительной степени компенсировавшую ее уменьшение от гибели во время загрязнения.[ ...]
Нектон — совокупность пелагических активно передвигающихся животных, не имеющих непосредственный связи с дном (киты, кальмары, ластоногие, рыбы).[ ...]
Толстолобики — крупные пелагические пресноводные рыбы, масса которых достигает 16 кг, длина — 1 м. Половой зрелости достигают в 5—6-летнем возрасте. Нерест — во время летнего паводка при температуре воды более 20 °С. Плодовитость — 490—540 тыс. икринок.[ ...]
А — Е — биотопные поля; 1 — пелагические рыбы, 2— придонные рыбы.[ ...]
Интервал между общим числом пелагических эхоотметок и их количеством в открытой части полигона II меньше, чем на предыдущем. Четко проявляется июльско-августовский пик численности мигрантов. Общая кривая динамики численности рыбного населения очень похожа на описанную для полигона I, но она сглажена и имеет более высокий зимний пик. Среднегодовой уровень численности рыб на полигоне II 55,2 шт./га, т. е. почти в 2 раза выше, чем в р. Паране.[ ...]
Подобно высшим позвоночным, рыбы могут широко варьировать частоту и амплитуду дыхания и ритм работы сердца в соответствии со своими энергетическими потребностями в той или иной ситуации и резервными возможностями. Вместе с тем для рыб характерно чрезвычайное разнообразие форм организации и функционирования аппарата внешнего дыхания [165, 422, 561]. Так, например, некоторым морским пелагическим видам (скумбрии, макрели, тунцы) и высокоподвижным пресноводным видам (язь) в норме свойственна пассивная перфузия жабр за счет встречного тока воды [53, 569]. Возможно, в естественных условиях всем реофилам в той или иной мере присуща "напорная" вентиляция жабр, а фиксированное состояние рыб, имеющее обычно место в экспериментальных условиях, сопряжено с дополнительной нагрузкой на респираторный аппарат. Есть виды рыб, утратившие способность активно вентилировать жабры во взрослом состоянии и вынужденные непрерывно плавать [569]. Характерно, что ритмы работы сердца и дыхательного аппарата у таких рыб необычно высокие для соответствующей температуры акклимации, хотя их соотношение такое же, как и у большинства рыб, т. е. ритм дыхания выше ритма сердечных сокращений [119, 121]. Так, например, при 18-22°С у макрели частота дыхания и сердечных сокращений равна 124 и 100 в минуту соответственно, а у кефали - 148 и 105 в минуту [569, 975]. Для сравнения напомним, что у карпа, например, величины этих показателей при 15°С равны 40 и 15 в минуту [416].[ ...]
Всего зарегистрировано 56 видов рыб из 28 семейств. Наибольшим количеством видов были представлены семейства корюшковых (4), лососевых (5), керчаковых (6), стихеевых (6) и камбаловых (5). Всех этих рыб можно разделить на три группы: 1)11 видов проходных и полупроходных рыб (карповые, лососевые, корюшковые и др.); 2) 38 видов донных, придонных и пелагических “местных” видов, всю жизнь проводящих в море и не совершающих дальних миграций; 3) 7 видов южных мигрантов (японский анчоус Engraulis japonicus, лобан Mugil cephalus, сардина иваси Sar-dinops melanostictus, сайра Cololabis saira, японская скумбрия Scomber japonicus, желтохвостая лакедра Serióla quinqueradiata, белоточечная собака-рыба Takifugu niphobles).[ ...]
На рис. 7.6 показана блок-схема модели сообщества пелагических рыб оз. Дальнего с учетом деления популяций на возрастные и половые группы. Озерная часть популяции красной представлена 4 возрастными группами: 0+, 1 + , 2+ и 3+, без разделения по половому составу. Таким образом, состояние озерной части популяции красной определяется 8 числами — численностями (Nf , . . ., N ) и средними весами (wfi . . . w%).[ ...]
В конце мая—начале июня 1985 г. основные скопления рыб располагались в средней части полигона, в русловой и прирусловой зонах (рис. 26). В верхней северной части полигона встречались отдельные обычно небольшие по размерам стаи рыб. По гидрофизическим показателям на полигоне выделялись две водные массы: стоковые воды Моложского плёса и воды Центрального плеса. Несколько различались они и по температуре. Эти различия (в пределах 0,5—1,0 °С) в весеннее время обусловлены более поздним таянием льдов в северной части полигона (Ривьер, 1988) Южная часть полигона мелководна: за пределами русла Мологи глубины обычно менее 3 м. Поэтому почти вся толща воды в этом месте подвергается ветровому воздействию, что исключает здесь длительное пребывание крупных скоплений пелагических рыб.[ ...]
Сообщество пелагических рыб озера Дальнего (опыт кибернетического моделирования). Наука», Л.[ ...]
Построение математических моделей популяций промысловых рыб и других водных животных является необходимым условием для эффективного управления биологическими ресурсами водоемов. В предлагаемой монографии изложены теоретические основы построения математических моделей популяций и сообществ водных животных. Основной метод моделирования — построение моделирующего алгоритма, реализованного на цифровой электронной вычислительной машине. На примере целого ряда абстрактных моделей поясняются принципы моделирования и устанавливаются некоторые закономерности динамики популяций. Переход к конкретным моделям осуществлен для популяции окуня в оз. Тюленьем и красной в р. Озерной. Специальная глава посвящена моделированию популяций водных беспозвоночных — планктонных ракообразных, причем рассматриваются и стохастические модели. Построение моделей сообществ начинается с абстрактных моделей и завершается конкретными моделями взаимодействия молоди окуня с диаптомусом в оз. Раздельном и сообщества пелагических рыб оз.[ ...]
| Блок-схема моделирующего алгоритма модели сообщества пелагических рыб оз. Дальнего. | 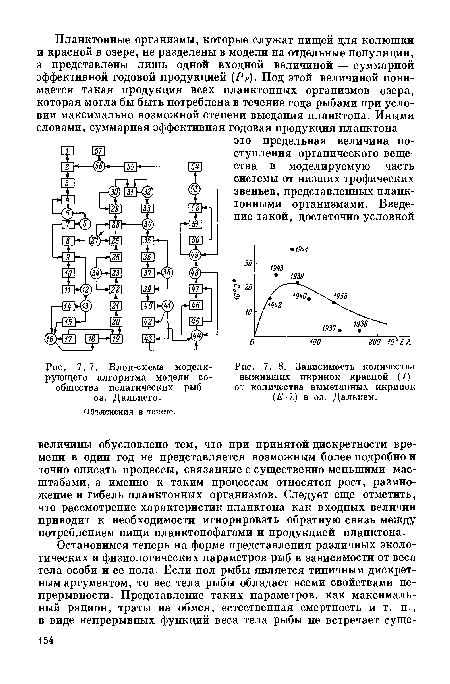 |
На рис. 28 представлено распределение наиболее плотных скоплений пелагических рыб по 2 горизонтам — 3—5 и 5—7 м. Видно, что зоны наибольшей плотности пелагических рыб совпадают, обычно в местах со сложным рельефом дна, а именно вблизи резких поворотов русел, их излучин. На ровных пойменных участках количество совпадений максимальных плотностей рыб в обоих горизонтах значительно меньше.[ ...]
| Связь в распределении в Тихом океане фосфатов, планктона и уловов пелагических рыб (по Богорову) | 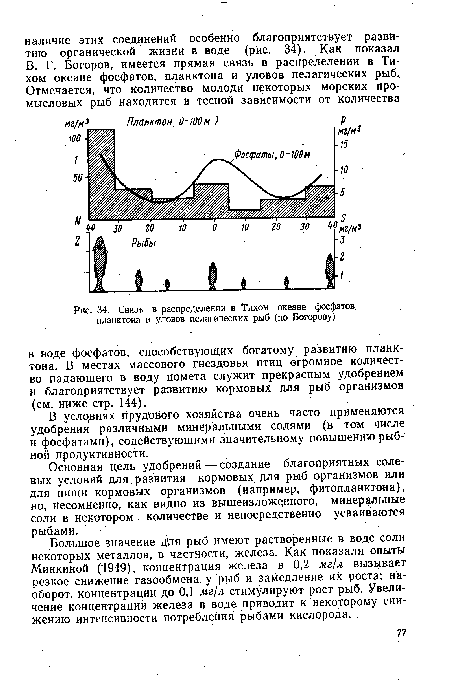 |
Наиболее выразительными представителями ископаемой макрофауны являются пелагические рыбы и их детрит, которые обычно избирательно концентрируются в определенных горизонтах (рис. 11.31). В алжирских диатомитах часто встречаются двустворчатые моллюски таких родов, как Peden, Cardita, Arca, Modiola, попадаются гастроподы, морские ежи, мшанки, брахиопо-ды, остракоды, а также листья и куски древесины.[ ...]
Пассивная нагульная миграция икры и личинок наблюдается также и у морских рыб. Пелагическая икра и личинки многих рыб переносятся течениями иногда на очень большие расстояния. Икра и личинки трески и личинки сельди переносятся атлантическим течением от берегов Норвегии к местам кормежки в Баренцевом море. Пелагическая икра многих камбал, сконцентрированная сразу после нереста на сравнительно ограниченной площади, постепенно течениями разносится на значительное пространство, чем обеспечиваются лучшие условия питания для выводящихся из икры личинок.[ ...]
Обращает на себя внимание увеличение в ихтиомассе доли обитающих в толще воды рыб с одновременным уменьшением придонных и донных особей. Если средняя биомасса рыб пелагического комплекса в первые 3 года (1978, 1979, 1982 гг.) была 75 кг/га, то в последующие 4 года (т. е. 1984, 1985, 1987, 1988 гг.) она возросла до 96 кг/га, а биомасса рыб донного комплекса уменьшилась соответственно с 59 до 41 кг/га. Это может быть связано с увеличением численности зоопланктона по всей толще воды и подъемом сюда значительной массы крупного половозрелого синца, ранее предпочитавшего придонные слои.[ ...]
О разногласиях и трудностях в систематике бычков указывается и в определителе рыб Черного моря А. Н. Световидова [1964]. Яот что он пишет по этому вопросу: «...относительно системы рода Gobius единого мнения нет. Ильин [1927, 1949] разделяет его на два рода (Gobius L. и Zostcrricola Iljin — Zosterissor whitiey), Берг [1949]—на три рода (Gobius, Neogobius Iljin и Mesogo bi-us), Крыжановский и Пчелина [1941] —также на три (Gobius, Eichwaldia Smitt — Hichwaldilla Whitiey и Mesogobius)» и далее: «...история происхождения и развития в этом вопросе, безусловно, имеет важное значение, однако разделение видов Gobius s. !. на упомянутые выше роды и отнесение их последними авторами к разным подсемействам и более крупным подразделениям „группам“, носят, по мнению Световидова, искусственный характер. Различаются эти роды и даже группы лишь по наличию или отсутствию плавательного пузыря, размерам икры и наличию или отсутствию пелагических личинок и определять их практически невозможно» [1964, с. 424—425].[ ...]
Суммарные данные по траловым уловам дают общую картину количественного распределения рыб в толще воды. Над глубинами более 12 м 63 % численности всех выловленных рыб приходится на горизонт 10—12 м, из них более 80 % составляет ряпушка, 23 % сосредоточено в горизонте от 2 до 4 м и представлено главным образом снетком. В среднем горизонте (5—7 м) отловлено всего 14 % рыб также с преобладанием снетка. На участках полигона, где глубины не превышают 10 м пелагические виды сосредоточены в слое воды 2—6 м и на 89 % состоят из снетка.[ ...]
Ихтиофауна оз. Модель сообщества пелагических рыб оз. Дальнего была выполнена в первом (Karpov a. oth., 1967; Карпов и др., 1969) и втором (Крогиус, Крохин, Меншуткин, 1969) приближениях. В настоящем разделе описывается только модель второго приближения.[ ...]
Вышедшая из икринки личинка имеет специальные приспособления для удержания в том местообитании, которое является для нее наиболее благоприятным, а именно: в том или ином слое воды (у пелагофильных рыб), в прибрежной зоне (для рыб литорали) и т. д. Личинки и мальки, ведущие пелагический образ жизни, очень часто имеют различные выросты, увеличивающие их сопротивление движению воды и облегчающие паре-! ние в ее Толще (см. выше). У фитофильных личинок рыб (сазан, лещ и многие другие), икра которых развивается в прикрепленном состоянии среди зарослей прибрежной зоны, часто имеется «цементный орган», пользуясь которым эмбрионы прикрепляются к растениям и другим предметам, находящимся в воде, что позволяет эмбрионам удерживаться в поверхностных слоях воды, „богатых кислородом.[ ...]
На рыбоводных заводах, где выращиваются мальки осетро вых, известны случаи, когда циклопы, помещаемые в бассейны как пища. осетрят, объедали у них мягкую часть плавников.[ ...]
Известна, например, чрезвычайно высокая плодовитость паразитов со сложным циклом развития; у таких животных вероятность прохождения отдельной особью полного цикла развития ничтожно мала, что эволюционно скомпенсировано высокой нормой плодовитости. Рыбы с пелагической икрой, доступной для выедания многими животными, откладывают ее в огромном количестве (например, кладка луны-рыбы Mota mola содержит до 300 миллионов икринок), тогда как виды, каким-либо способом укрывающие икру, характеризуются более низкой плодовитостью (тысячи и сотни икринок на самку), а обладающие активными формами заботы о потомстве продуцируют небольшие порции икры. Акулы, яйца которых защищены плотной оболочкой, откладывают всего несколько яиц в сезон. Среди бесхвостых амфибий не обладающая заботой о потомстве зеленая жаба Bufo viridis производит за сезон 8—12 тысяч икринок, тогда как жаба-повитуха Afytes obsíeíri-cans, наматывающая икру на задние лапы,— всего 150, а пипа Pipa americana, вынашивающая ее в особых ячейках в коже спины,— порядка 100 икринок.[ ...]
Начальной стадией работ по мониторингу загрязнения морских гидро-бионтов бенз(а)пиреном явилась оценка влияния условий хранения отобранных проб в полевых условиях (на судне) на результаты исследований. Объектами исследований были наиболее массовые промысловые виды рыб юго-восточной части Баренцева моря, отличающиеся структурой мышечной ткани и жирностью - пелагические виды (сельдь, корюшка) и донные виды (навага), отобранные в относительно чистом - Поморском проливе и загрязненном районе - район Варандея.[ ...]
Другая важнейшая адаптивная реакция, возникающая при длительном или краткосрочном дефиците кислорода в окружающей среде, но уже на биохимическом (молекулярном) уровне — это изменение сродства гемоглобина к кислороду. Еще в начале текущего столетия А. Крог и И. Лейч [469] показали, что адаптация рыб к пониженному содержанию кислорода осуществляется за счет повышения сродства гемоглобина к кислороду. Сопоставляя величину напряжения кислорода в воде, необходимую для полунасыщения крови у малоподвижных пресноводных рыб (карп, угорь), часто встречающихся с дефицитом кислорода в естественных условиях обитания, с высокоподвижной оксифильной форелью, они обнаружили, что у малоподвижных рыб эта величина в 3—5 раз ниже, чем у высокоподвижных. Такая же зависимость выявлена и при сопоставлении двух, различающихся по уровню активности, видов морских рыб — придонной камбалы и пелагической трески, однако в этом случае различия достигали лишь двукратной величины (рис. 18) ■ Исследования этого плана были продолжены на морских рыбах Р. Рутом [572], который пришел к выводу, что кровь высокоактивных рыб обладает повышенной кислородной емкостью в сравнении с кровью малоактивных рыб. По мнению ряда специалистов [77, 79, 368], степень сродства гемоглобина к кислороду является важнейшим фактором, определяющим уровень устойчивости рыб к дефициту кислорода. Выявлено наличие связи между величинами Р о и Р95 крови и уровнем пороговых и критических /э02 (рис. 19) для многих морских и пресноводных видов рыб, относящихся к различным по уровню активности экологическим группам [79] .[ ...]
С увеличением интенсивности прибрежного промысла ихтио-масса молоди красной в озере сначала медленно повышается, при Р=0Л достигает максимума, а затем начинает резко сокращаться (рис. 7.14). То же можно сказать и об ихтиомассе популяции гольца. Характерно, что при полном уничтожении популяции красной ( 0.8) гибнет и популяция гольца — она оказывается неспособной существовать, используя в качестве источника питания одну лишь колюшку. Этот результат — не отражение в модели реальных свойств сообщества пелагических рыб оз. Дальнего, а следствие недостаточно подробного описания в модели приспособительных свойств популяции гольцов.[ ...]
Жизнь в донном районе резко отличается по своим условиям от жизни в прибрежной полосе. Здесь при большом давлении в десятки атмосфер (на каждые 10 метров глубины давление увеличивается на одну атмосферу) царствует полная неподвижность окружающей среды, однообразная низкая температура круглый год и полная или почти полная темнота. Животные на больших глубинах бесцветны или слабо окрашены в дополнительный розовый, красный цвет, часто с редуцированными глазами, но с развитыми органами осязания. Кроме того, в тропических и притропических морях встречаются рыбы и головоногие моллюски с глазоподобными органами, но испускающими свет вследствие сгорания вырабатываемого ими особого вещества (фотоген). Впрочем, эта способность свечения прйсуща очень многим животным всрх типов, живущим в совершенно других условиях в третьем районе — пелагическом. В пресных водах из рыб для пелагического района характерными являются: уклея, снеток, ряпушка, вообще сиги; но здесь же живет и молодь некоторых прибрежных рыб, напр, леща, на некоторое время плотвы, окуня, язя и других; в морской—все виды сельдей, макрель. Все формы постоянно пелагических видов отличаются более или менее своей прозрачностью, доходящей до полной стекловидности, как, например, С1иреопе11а Сшппи Каспийского моря, 2 личинки салаки в Балтийском и хамсы — в Черном море.[ ...]