Хищники же, скорее всего, следуют за колебаниями численности зайца, а не вызывают их. Все же колебания, вероятно, выражены более отчетливо благодаря высокому отношению числа хищников к числу жертв в период снижения численности зайца, а также благодаря их низкому отношению в период, следующий за минимумом численности зайцев, когда они, опережая хищника, восстанавливают свою численность (рис. 10.5). Кроме того, при высоком отношении численности рыси к численности зайца хищник поедает большое количество боровой дичи, а при низком отношении — небольшое. Это, по-видимому, служит причиной возникновения колебаний численности у этих второстепенных растительноядных животных (рис. 10.5). Таким образом, взаимодействие зайцы—растения вызывает колебания численности зайца, хищники повторяют колебания их численности, а циклы численности у растительноядных птиц вызваны изменениями пресса хищников. Очевидно, что простые модели полезны для понимания механизмов колебаний численности в природных условиях, но эти модели объясняют возникновение этих колебаний далеко не полностью.[ ...]
Высокая численность вида-прокормителя создает благоприятные условия для размножения питающихся им хищников, паразитов и возбудителей болезней. Есть виды, для которых хищники — главный регулятор численности: если жертв много, хищники хорошо питаются, быстро размножаются и пожирают все большую часть жертв, но, снизив численность жертв, сами резко уменьшаются в численности. При низком уровне численности хищников жертвы вновь размножаются, вслед за чем повышается численность жертв, — и цикл повторяется вновь. Те, кто скупает меха, давно заметили явление колебаний численности у пушных зверей. Классическим примером являются колебания численности зайца-беляка и рыси.[ ...]
Присущее взаимодействие типа хищник—жертва свойство порождать взаимосвязанные колебания численности вызывало иногда в прошлом «надежды» на то, что такие же колебания будут обнаружены в реальных популяциях. Эти надежды, однако, следует сразу же умерить по двум соображениям. Во-первых, существует много важных аспектов экологии хищника и жертвы, которые в разработанных моделях пока не учтены; однако, как мы увидим в следующих разделах, эти аспекты могут существенно изменить любые прогнозы. Во-вторых, даже если в популяции наблюдаются регулярные колебания численности, то это совсем не обязательно служит подтверждением модели Лотки—Вольтерры, логистической модели с запаздыванием или любой другой простой модели. Если численность популяции растительноядного животного колеблется, то это может отражать ее взаимодействия с пищевыми объектами или с хищниками. И если численность популяции жертвы циклически меняется в силу присущих ей свойств, то численность хищников может повторять эти циклы, даже если само взаимодействие хищник—жертва их не вызывает. Таким образом, взаимодействия типа хищник—жертва могут порождать регулярные колебания численности обеих взаимодействующих популяций и могут усиливать такие колебания, если они существуют вследствие других причин; но выявление причины регулярных циклических колебаний в естественных условиях представляет собой, как правило, трудную задачу (см. разд. 15.4).[ ...]
Таким образом, при хорошей адаптации хищника в окрестности устойчивого равновесия могут возникать неустойчивый и устойчивый циклы, т.е. в зависимости от начальных условий система ’’хищник—жертва” либо стремится к равновесию, либо, колеблясь, уходит от него, либо в окрестности равновесия устанавливаются устойчивые колебания численностей обоих видов.[ ...]
Если равновесие неустойчиво (предельных циклов нет) или внешний цикл неустойчив, то численности обоих видов, испытывая сильные колебания, уходят из окрестности равновесия. Причем быстрое вырождение (в первой ситуации) наступает при низкой адаптации хищника, т.е. при его высокой смертности (по сравнению со скоростью размножения жертвы). Это означает, что слабый во всех отношениях хищник не способствует стабилизации системы и сам вымирает.[ ...]
| Модель Лотки—Вольтерры для системы хищник—жертва. А. Изоклина для популяции жертвы. При низкой плотности хищника (С) численность жертвы (Л/) возрастает, а при более высокой — снижается. Б. Изоклина для •популяции хищника. При высокой плотности жертвы численность популяции хищника растет, а при низкой — снижается. В. При объединении изоклин в системе хищник—жертва возникают неограниченные взаимосвязанные колебания численности, подобные тем, которые показаны на рис. Г. Однако, как видно на рис. Д, для этих колебаний характерна нейтральная стабильность | 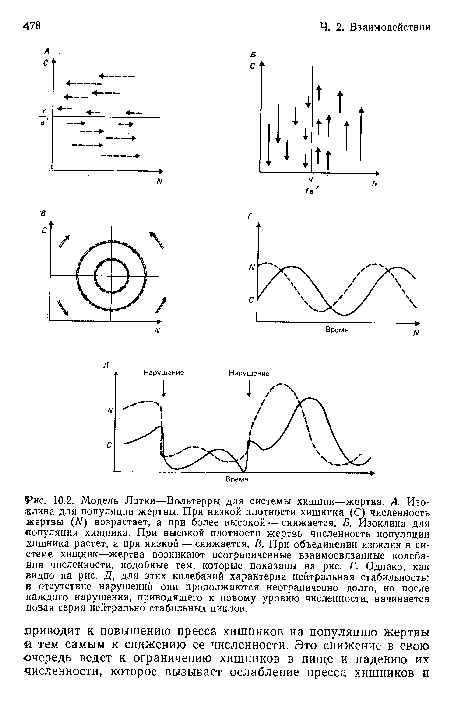 |
Теории четвертой группы основаны на представлении, согласно которому регуляция циклов численности осуществляется на уровне экосистемы, а не на уровне популяции. Разумеется, изменения плотности, охватывающие диапазон в несколько порядков величины, должны сказываться не только на вторичном трофическом уровне (хищники и жертвы), но также на первичном звене трофической цепи — растениях и травоядных животных. Рассмотрим в качестве примера «гипотезу возврата питательных веществ» Шульца (1964, 1969), предложенную для объяснения небольших колебаний плотности популяций тундры. Со-гласно этой гипотезе, которая подтверждается данными исследований круговоротов неорганических веществ, в результате сильного объедания растительности леммингами в годы взрыва их численности в последующие годы происходит уменьшение доступных минеральных питательных веществ (особенно фосфора), и питательная ценность пищи леммингов сильно уменьшается. В связи с этим рост и выживание молодых особей значительно снижаются. В течение третьего и четвертого года питательные вещества возвращаются в цикл, растительный покров восстанавливается и экосистема вновь может обеспечить высокую плотность леммингов.[ ...]
Они не подразумевают стабильной популяции. Вместо этого они описывают циклические колебания численности хищника и жертвы (рис. 2-11). В каждом цикле, как только увеличивается популяция жертвы, начинает расти и популяция хищника. Она превышает численность жертвы, потребляет кормовой ресурс больше нормы для поддержания своей стабильности, отчего размер популяции жертвы снижается. Теперь численность хищника следует за убыванием численности жертвы до тех пор, пока уровень плотности хищника не станет таким низким, что популяция жертвы начинает вновь возрастать, и в дальнейшем хищник снова принимается следовать этому движению вверх. Если считать условия окружающей среды постоянными, тогда интенсивность, или ам-плитуда, циклов — разница между «пиком» и «дном» плотности — будет определяться начальной плотностью и останется неизменной. В этом случае популяции хищника и жертвы могут испытывать циклические колебания своей численности вечно.[ ...]
В природе происходят периодические изменения условий, в частности климатических, связанные с циклами солнечной активности, периодическими изменениями основных направлений переноса влаги ветрами и с другими причинами, часто взаимодействующими слишком сложным образом, чтобы можно было всегда уверенно указать основные причины подъема или снижения численности того или иного вида. Кроме того, многие виды зависят от численности друг друга, как, например, в случае взаимодействия хищников и жертв (рис. 2.14). Численность многих видов соответственно имеет 11-летние, связанные с солнечными, и другие циклы, в которых бывают минимумы, когда вид становится наиболее уязвимым. Это явление особенно характерно для относительно простых экосистем, где обычно небольшое число видов представлено большим числом особей. Для выживания в таких экосистемах с суровыми и часто переменчивыми условиями жизни оказалась, очевидно, выгодной и закрепилась эволюцией изменчивость численности в широких пределах как форма существования видов. Простейший пример такого вида с большой амплитудой колебаний численности представляет обыкновенная домашняя муха, численность которой в конце лета и ранней весной может отличаться в сотни тысяч раз.[ ...]
Существует ряд примеров, полученных на природных популяциях, в которых можно обнаружить регулярные колебания численности хищников и жертв. Колебания численности популяций зайца обсуждаются экологами, начиная с двадцатых годов нашего века, а охотники обнаружили их еще за 100 лет до того. Так например, американский заяц-беляк (Lepus americanus) в бореальных лесах Северной Америки имеет «10-летний цикл численности» (хотя на самом деле его продолжительность варьирует от 8 до 11 лет; рис.[ ...]
И, наконец, последняя устойчивая ситуация возникает при очень низких приспособленности и смертности хищника. Здесь устойчивый цикл лишь ’’размывается” случайными возмущениями, что приводит к флуктуации в периодических колебаниях численности хищника и жертвы.[ ...]
В. Вольтерры, как уже упомянуто ранее, предложенные ими независимо друг от друга в 1925 и 1926—1931 гг. На эти уравнения буквально набросились прикладные математики экологического направления. Они породили огромную литературу. Еще в начале 30-х гг. выраженная ими закономерность была экспериментально проверена Г. Ф. Гаузе (1934), получившего опытные доказательства справедливости уравнения А. Лотки — В. Вольтерры. Последний сформулировал три закона системы «хищник — жертва». Закон периодического цикла: процесс уничтожения жертвы хищником нередко приводит к периодическим колебаниям численности популяций обоих видов, зависящим только от скорости роста популяций хищника и жертвы и от исходного соотношения их численностей. Закон сохранения средних величин , средняя численность популяции для каждого вида постоянна независимо от начального уровня при условии, что специфические скорости увеличения численности популяций, а также эффективность хищничества постоянны. Закон нарушения средних величин: при аналогичном нарушении популяций хищника и «ертвы (например, рыб в ходе промысла пропорционально их численности) средняя численность популяции жертвы растет, а популяции хищника падает.[ ...]