КЛИМАКС КЛИМАТИЧЕСКИЙ (региональный) —стабильное состояние сообщества (экосистемы), находящегося в равновесии с общими климатическими условиями [55].[ ...]
Климакс, климакс-формация стабильное состояние биоценоза, достигнутое в результате развития сообщества, завершения сукцессии.[ ...]
Климакс — завершающая стадия сукцессии для данных условий среды.[ ...]
Климакс — финальная относительно устойчивая фаза естественного развития биогеоценоза или растительного сообщества, наиболее соответствующая экологическим характеристикам данной местности в определенный период геологического времени.[ ...]
Климакс—относительно стабильное состояние биоценоза (экосистемы).[ ...]
КЛИМАКС [гр. кИтах — высшая точка, кульминация] — стабильное состояние сообщества (экосистемы), в котором климаксовое сообщество (экосистема) поддерживает само себя неопределенно долго, все внутренние его компоненты уравновешены друг с другом.[ ...]
Достижение климакса при потере одного или группы видов в результате их уничтожения (реже вымирания, тоже, как правило, связанного с антропогенным исчезновением местообитаний) не есть полное восстановление природной обстановки. Фактически это новая экосистема, так как в ней возникли новые связи и утеряны многие старые, сложилась иная «при-тертость» видов. Вернуться в старое состояние экосистема не может, так как утерянный вид невосстановим. Это позволяет сформулировать закон эволюционно-экологической необратимости: экосистема, потерявшая часть своих элементов или сменившаяся другой в результате дисбаланса экологических компонентов, не может вернуться к первоначальному своему состоянию в ходе сукцессии, если в ходе изменений произошли эволюционные перемены в экологических элементах (сохранившихся или временно утерянных) или один либо группа видов исчезла навсегда или на (системно) долгий срок. Если какие-то виды утеряны в промежуточных фазах сукцессии, то эта потеря может быть функционально скомпенсирована, но лишь частично. При снижении разнообразия за критический уровень ход сукцессии искажается и фактически климакс, идентичный прошлому, достигнут быть не может.[ ...]
Сукцессия и климакс экосистем. Изменение внешних условий среды влияет на некоторые виды неблагоприятно, другие же виды могут от этого, наоборот, выиграть. Подчас изменившиеся условия позволяют включиться в экосистему новым видам. В целом происходит так называемая сукцессия (от лат. зиссевю — преемственность): последовательная необратимая смена биоценозов, преемственно возникающих на одной и той же территории в результате влияния природных факторов или воздействия человека.[ ...]
У концепции климакса долгая история. Один из первых исследователей сукцессий Фредерик Клементс (Clements, 1916) утверждал, что в любой климатической зоне существует только один истинный климакс. К его возникновению ведут все сукцессии независимо от того, начались ли они с песчаной дюны, залежи или даже зарастающего пруда, постепенно превращающегося в наземное сообщество. Такую крайнюю теорию моноклимакса многие экологи, среди которых и такой выдающийся как Тенсли (Tansley, 1939), в конце концов отвергли. Школа поликлимакса признавала, что климакс в данном участке может определяться одним или несколькими факторами: климатом, почвенными условиями, топографией, пожарами и т. д., поэтому в одной климатической зоне вполне может существовать целый ряд специфических типов климакса. Еще позднее Уиттекер (Whittaker, 1953) предложил свою гипотезу климаксовой структуры пространства. По его мнению в природе существует континуум типов климакса, постепенно меняющихся по средовым градиентам и совсем необязательно разделенных четкими границами (это является экстраполяцией его подхода к градиентному анализу, рассмотренного в разд. 16.3.1).[ ...]
Климатический климакс является результатом длительной сук-цессионной серии экосистемы в данных климатических условиях. Но, как известно, для Земли характерна климатическая зональность, а отсюда и зональность наземных экосистем, климаксная стадия которых будет определяться конкретными климатическими факторами соответствующей зоны. Известно, что кроме горизонтальной зональности (рис. 7.4) климата в масштабе всего земного шара, наблюдается еще и вертикальная или высотная зональность в горных системах. Югом» будет являться климат подножия горной системы (см. рис. 7.4).[ ...]
Климатический климакс является результатом длительной сук-цессионной серии экосистемы в данных климатических условиях. Но, как известно, для Земли характерна климатическая зональность, а отсюда и зональность наземных экосистем, климаксная стадия которых будет определяться конкретными климатическими факторами соответствующей зоны. Известно, что кроме горизонтальной зональности (рис. 7.4) климата в масштабе всего земного шара, наблюдается еще и вертикальная или высотная зональность в горных системах. Югом» будет являться климат подножия горной системы (см. рис. 7.4).[ ...]
| Эдафические климаксы на прибрежном участке в Северной Калифорнии (по |  |
С общеэкологических позиций климакс рассматривается как динамически равновесное состояние сообщества, обеспечивающее постоянство видового состава и поддерживающее в целом устойчивое структурное разнообразие элементов за счет внутриценотических нарушений разного масштаба и разной природы. С популяционных позиций климакс характеризуется устойчивыми потоками поколений в популяциях всех потенциальных обитателей данной территории.[ ...]
Снижение разнообразия видов в климаксе не означает малой его экологической значимости. Разнообразие видов формирует сукцессию, ее направление, обеспечивает заполненность реального пространства жизнью. Недостаточное количество видов, составляющих комплекс, не могло бы сформировать сукцессионный ряд, и постепенно, с разрушением климаксовых экосистем произошло бы полное опустынивание планеты. Значение разнообразия функционально как в статике, так и в динамике. Следует отметить, что там, где разнообразие видов недостаточно для формирования биосферы, служащей основой нормального естественного хода сукцессионного процесса, а сама среда резко нарушена, сукцессия не достигает фазы климакса, а заканчивается узловым сообществом — параклимаксом, длительно или кратковременно производным сообществом. Чем глубже нарушенность среды того или иного пространства, тем на более ранних фазах оканчивается сукцессия.[ ...]
Снижение разнообразия видов в климаксе на первый взгляд противоречит рассмотренному стремлению к биоразнообразию в природе. Однако именно разнообразие видов формирует сукцессию и ее направление, обеспечивает заполнение реального пространства жизнью.[ ...]
| Климатический и эдафический климаксы на востоке США. |  |
На одном прибрежном участке северной части Калифорнии лес из гигантских секвой растет бок о бок с карликовым лесом из крошечных, чахлых деревьев. Ка« видно на схеме, один и тот же песчаник подстилает эти леса, но карликовый лес встречается там, где водонепроницаемый горизонт залегает близко к поверхности, сильно ограничивая рост корней, а также движение воды и питательных веществ. Растительность, достигшая равновесных условий, или климакса, в этой частной ситуации, почти полностью отличается по видовому составу и структуре от растительности прилежащих районов, где водонепроницаемый горизонт либо отсутствует, либо залегает глубоко.[ ...]
Чтобы лес восстанавливался до состояния климакса, требуется длительный суккцессионный цикл. Для ускорения процесса предлагается, например, вырубать его узкими просеками, оставляя растения, которые ценности для промышленности не представляют, не нарушая при этом запас биогенов в корневых подушках, и тогда обсеменение с незатронутых участков поможет быстро восстановить лес до первоначального вида.[ ...]
Существует еще такое понятие, как катастрофический (или циклический) климакс. Катастрофический климакс - терминальное состояние экосистемы, возникающее благодаря периодически появляющимся катастрофам, таким как пожары, вредители и т. д. Природные катастрофы уничтожают климаксное сообщество и способствуют тем самым повторению сукцессии, длящейся до климаксного состояния, при котором экосистема особенно подвержена воздействию катастрофического фактора. Подобный процесс повторяется неоднократно, и со временем возникает естественная самоподдер-живающаяся система, для существования которой необходимы регулярно возникающие катастрофы.[ ...]
Анализ материалов позволил установить, что развитие перифитона до стадии климакса занимает около 50 дней, при этом сукцессия сообщества обрастателей состоит из ряда этапов. Стадия колонизации продолжается 1-2 сут.; для нее характерно преобладание процессов заселения субстрата из толщи воды (0.030 мг/дм2 в сут.) над размножением водорослей (0.017 мг/дм2 в сут.). Таксономическая структура сообщества в это время отражает структуру планктоценозов (доминирование в биомассе планктонных Diatoma, Frag Haría, Melosira, крайне низкая роль прикрепленных водорослей базального слоя Achnanthes и Cocconeis). Вторая стадия -формирование базального слоя - продолжается около 15 дней. Для ее начала характерно доминирование Achnanthes (23.7-44.8%) при большой роли Cocconeis (17.3-20.0%), в конце стадии доминируют Coccone/s(32.1%) при высокой доле Achnanthes (16.9%), биомасса сообщества достигает 12.4-15.7 мг/дм2. В третьей стадии (продолжительность 10-20 дней) образуется двуслойное сообщество, в котором доминируют Gomphonema olivacea (8.7-17.8%), Cymbeiia cistula (14.6-15.1%), виды рода Navícula (16.2-27.4%), его биомасса достигает 31.1-42.7 мг/дм2. В четвертой, завершающей стадии формируется многослойный перифитический мат; на первый план выдвигаются крупные водоросли дистального слоя Synedra ulna (25.8-32.7%) и Cymbella lanceo lata (18.8-39.7%). В стадии климакса наблюдаются нерегулярные колебания биомассы водорослей (110.3-223.9 мг/дм2), связанные с периодическим отслаиванием части перифитонных матов, при неизменно высокой роли S. ulna и C. lanceolata.[ ...]
Соотношение семейств пауков варьирует в широких пределах. По мере приближения к климаксу наблюдается сопряженное изменение попадае-мости наиболее обильных семейств Lycosidae и Gnaphosidae. Обильные виды отсутствуют на ранних стадиях сукцессии. В климаксе содоминируют два вида - Trochosa terrícola Thorell,1856 и Zelotes subterraneus (C.L. Koch, 1833).[ ...]
Биом включает в себя не только климатическую климаксную рас-титёльность, но и эдафические климаксы и стадии развития, в которых во многих случаях доминируют иные жизненные формы (гл. 9). Так, травянистые сообщества .представляют собой временную стадию развития в биоме листопадного леса, где климаксной жизненной формой; являются широколиственные листопадные деревья. Биом идентичен главной «растительной формации» в том смысле, в каком этот термин используется в экологии растений; разница лишь в том, что биом — это единица всего сообщества, а не одной лишь растительности. В биом входят не только растения, но и животные. В общем можно сказать, что биом соответствует «главной биотической зоне», если понимать под этим выражением зону сообщества, а не флористическую или фа-унистическую единицу. Биом соответствует «главной зоне жизни» в понимании европейских экологов, но не соответствует «зоне жизни», как ее понимают американские экологи. В Северной Америке термин «зона жизни» относят к ряду «температурных зон», описанному в 1894 г. К- Мерриэмом и широко используемому при изучении птиц и млекопитающих. Первоначальные температурные критерии были отброшены, и в .настоящее время в основу мерриэмовских зон жизни положено распределение животных, так что эти зоны все более и более превращаются в зоны сообществ и во многих случаях это то же самое, что и части и подсистемы биомов. Сравнение биомов и зон жизни Мерриэма см. у Ю. Одума (1945).[ ...]
В лесоведении прочно удерживается представление о старых вполне сформировавшихся, достигших стадии климакса лесах не как о биогеоценозах, наиболее гармонично соответствующих условиям данной природной зоны и ее климату, а как о перестойных лесах. Иначе, как о лесах, которые превысили срок, когда древостой их достигал биологической и технической спелости. Отсюда вывод, что если такие леса будут существовать далее, то экономическая их ценность снизится, они перезреют, «перестоят». Это экономическая, технологическая сторона вопроса, и она вполне правомочна. Другая сторона этой теории — биологическая, а точнее биоценологическая. Она состоит в утверждении, что такое насаждение, предоставленное самому себе, будет охвачено процессом распада, произойдет саморазрушение лесного насаждения. Это положение глубоко порочно: естественный процесс возрастной эволюции не может вести к необратимому процессу смерти, распада, разрушения лесной ассоциации, биогеоценоза как такового.[ ...]
Сукцессия — последовательная смена одних биогеоценозов другими. Конечным результатом сукцессии является климакс — финальная, относительно устойчивая фаза развития БГЦ.[ ...]
При потере одного или группы видов в результате их уничтожения (антропогенное исчезновение местообитаний, реже вымирание) достижение климакса не является полным восстановлением природной обстановки. Фактически это новая экосистема, потому что в ней возникли новые связи, утеряны многие старые, сложилась иная «притер-тость» видов. В старое состояние экосистема вернуться не может, так как утерянный вид восстановить невозможно.[ ...]
Биогеоценоз коренной — соответствующий географическим (природно-антропогенным) условиям местности и иочти не нарушенный человеческой деятельностью (см. климакс сукцессиоииый).[ ...]
Человек часто влияет на развитие экосистемы, препятствуя достижению ею климаксного состояния. Когда сообщество, не представляющее собой климатический или эдафический климакс для данной местности, поддерживается человеком или домашними животными, то его называют дисклимаксом, или антропогенным субкпимаксом. Например, чрезмерный выпас может породить пустынное сообщество там, где по условиям регионального климата могла бы сохраняться степь. Пустынные сообщества в данном случае - дисклимакс, а степь - климатический климакс.[ ...]
Сукцессия завершается формированием сообщества, наиболее адаптированного по отношению к комплексу сложившихся климатических условий. Концепция «климакса» подразумевает, что в пределах региона с более или менее однородным климатом фитоценозы, завершившие сукцессионный процесс, образуют климаксные сообщества независимо от того, с какого типа начиналась сукцессия.[ ...]
Сукцессия, в энергетическом смысле, связана с фундаментальным сдвигом потока энергии в сторону увеличения количества энергии, направленной на поддержание системы. Стадии роста, стабилизации и климакса сукцессии можно различать на основе критерия продуктивности: на первой стадии продукция растет до максимума, на второй остается постоянной, а на третьей по мере деградации системы уменьшается до нуля.[ ...]
Возможна и третья точка зрения. Ненарушенные сообщества обычно переходят одно в другое по градиентам местообитаний. Климаксовая растительность, таким образом, не состоит из какого-то определенного числа четко отличимых типов климакса. Мы можем рассматривать третью точку зрения на климаксовую растительность как концепцию климакса-континуума. Климакс — это сообщество в устойчивом состоянии, признаки которого определяются свойствами его местообитания, или биотопа. Развивающиеся в этих условиях местообитания и сообщества переходят в другие, образуя экоклины. Несмотря на случаи нарушений и дискретности, климаксовые сообщества следует интерпретировать не как мозаику отграниченных друг от друга типов, а как сочетание сообществ, переходящих одно в другое в соответствии с изменением градиентов условий внешней среды. Такое распределение включает ряд различных типов сообществ, приспособленных к ряду топографических и почвенных условий данного района. В этом разнообразии, однако, можно видеть центральный или наиболее распространенный (в устойчивом состоянии ненарушенный) тип сообществ, который вбирает в себя наибольшее число климаксовых сообществ района и который занимает большую часть местообитаний, не являющихся в данном регионе экстремальными или по каким-либо причинам специфическими. Этот тип сообществ есть превалирующий, или климатический, климакс, и его можно рассматривать как отражение климата районов.[ ...]
| Кривые динамики видового богатства для трех освобождающихся время от времени участков н сообщества в целом при различных частотах нарушения. Прерывистые линии соответствуют стадии конкурентного исключения перед наступлением климакса | 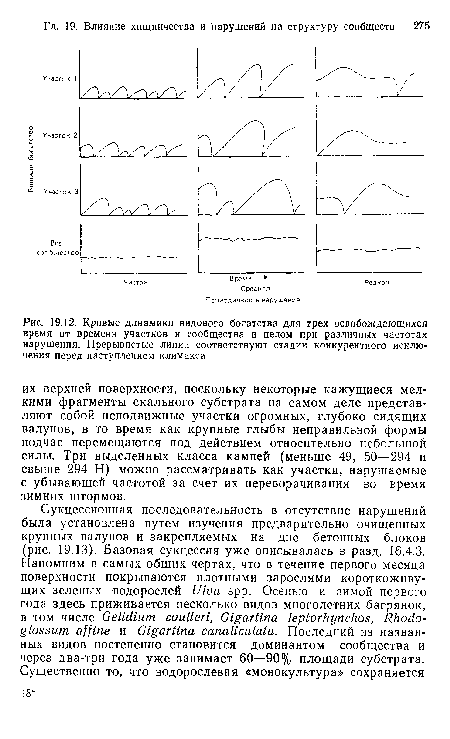 |
| 5.1 |  |
Представление о том, что автогенное развитие неизбежно приводит к стабильному сообществу, широко принято, поскольку оно исходит из глубоких наблюдений и здравой теории1. Однако по интерпретации этого представления экологи делятся на две школы. Согласно концепции «моноклимакса», в любой зоне возможен лишь один климакс, по направлению к которому развиваются, хотя и медленно, все сообщества. Согласно концепции «поликлимакса», нельзя считать реальным, что все сообщества в данной климатической зоне придут к одному и тому же, несмотря на все разнообразие физической среды. Нельзя также рассчитывать, что под влиянием данного сообщества все местообитания достигнут какого-то единого уровня за приемлемые отрезки времени, сопоставимые с продолжительностью жизни человека (или в несколько раз большими!). Удобный компромисс между этими двумя точками зрения — признание единственного теоретического климатического климакса и различного (в зависимости от изменчивости субстрата) числа эдафических климаксов. Таким образом, климатический климакс — это теоретическая константа, с которой можно сравнивать наблюдаемые условия. При этом становится возможным измерить степень отклонения от теоретического климакса, если оно наблюдается, и легче определить факторы, ответственные за это отклонение, поскольку имеется некий исходный «эталон» для сравнения. Одна из важных причин необходимости сохранить нетронутые уголки природы состоит в том, что для каждой обширной географической области следует иметь естественный климакс, с которым можно было бы сравнивать различные окультуренные ландшафты.[ ...]
Циклические сукцессии имеют длительный (десятки лет) период смены биоценозов на фоне динамики экотопа с возвратом к исходному состоянию по истечении определенного периода времени. В определенных условиях экосистема в процессе сукцессии может приходить к длительному под-вижно-стабильному состоянию, именуемому климаксом (от греч. климакс - лестница). Климаксными являются некоторые таежные экосистемы, целинные ковыльные степи.[ ...]
Действительно, в полевых условиях очень трудно выделить устойчивое климаксовое сообщество. Обычно удается лишь заметить, что скорость сукцессии падает до определенного уровня, после которого мы уже не видим никаких изменений. На залежах достижение «климаксовой» стадии потребовало бы 100—300 лет, однако вероятность возникновения пожара или урагана за этот период настолько высока (в Новой Англии ураганы случаются примерно раз в 70 лет), что сукцессия может никогда не завершиться.[ ...]
Другим физическим фактором, периодические воздействия которого имели жизненно важное значение для человека и природы на протяжении столетий, является огонь. На протяжении многих веков человек сознательно использовал пожары для поддержания таких климаксов или для возвращения сукцессии на какую-нибудь из ранних стадий.[ ...]
ВРЕМЯ ЭКОЛОГИЧЕСКОЕ КОРОТКОЕ — время перехода в обл. локального равновесия с максимумом локального минимума потенциальной энергии или максимумом локального минимума биологического разнообразия; приблизительно равно сумме времен релаксации при неравновесных переходах из обл. локального равновесия. Время сукцессий от полностью разрушенного до сообщества климатического климакса (от десятков для травяных сообществ до сотен лет для древесных). В общем случае зависит от взаимодействия с другими базовыми переменными и в т. ч. от пространства, для которого реализуется процесс: чем больше пространство, тем больше В.э.к. [76].[ ...]
Таким образом, конечная стадия восстановительных сукцессий (демутаций) в современном растительном покрове лесного пояса -разновозрастный лес с хорошо выраженной мозаикой окон возобновления и ветровально-почвенных комплексов. В зависимости от полноты представленности видов региональной флоры, способных существовать под пологом леса и в окнах, такой лес может быть определен как климакс или диаспорический субклимакс. Климаксным лесным сообществом мы будем называть разновозрастный лес с выраженной gap-мoзaикoй и с максимальным числом лесных видов региональной флоры. При ограниченных возможностях заноса зачатков лесных видов региональной флоры демутация приводит к формированию диаспорического субклимакса. Так же как и климакс, диаспорический субклимакс способен к длительному спонтанному существованию, однако он не содержит в своем составе многих видов, способных по своим экологическим свойствам обитать в лесных сообществах.[ ...]
Раздел 1. Общие закономерности организации биогеоценотического покрова. Сообщество как множество взаимодействующих популяций. Основные параметры элементарных популяций растений и животных. Популяционная организация биогеоценотического покрова лесных территорий. Представления о сукцессиях и климаксе с популяционных позиций.[ ...]
НАДЕЖНОСТЬ ЭКОЛОГИЧЕСКАЯ (экосистемы) — способность экосистемы относительно полно самовосстанавливаться и саморегулироваться (в пределах естественных для системы суточных, сезонных, межгодовых и вековых колебаний) в течение сукцессионного или эволюционного периода ее существования. Важной характеристикой экологической надежности служит сохранение структуры, функций и направления развития экосистемы, без которых данная экосистема сменяется другой, с иными функциями и структурой, а иногда и направлением развития к иной фазе климакса (любого типа) или узлового сообщества. Например, биогеоценозы кедровых лесов Алтая могут сменяться луговой растительностью или лиственничным лесом как конечными образованиями, кедр при этом не восстанавливается.[ ...]
Гидротехнические мелиорации изменяют не только гидрографию территории (создают новые водные объекты), но и меняют гидрогеологические условия (подъем или понижение уровня грунтовых вод, их засоление) и микроклимат (изменение амплитуды колебаний температур воздуха, его относительной влажности и др.). При этом резко преобразуются эдафические экологические условия — вторичное засоление, слитизация и деградация черноземов при орошении; ослабление процессов торфообразования и возникновение опасности дефляции торфяников при осушении и др. Это приводит к нарушению климаксов фитоценозов и возобновлению сукцессий биоты (отмирают одни древесные и травянистые растения и появляются другие, обедняется видовой состав растительных сообществ), вымирают или вытесняются отдельные виды животных при орошении земель, строительстве каналов, осушении болот, нарушениях мест обитания, разрыве трофических цепей, перекрытии путей миграции копытных и др.[ ...]
Б. Условия температуры и влажности в Тель-Авиве в разные годы по сравнению с оптимальными и благоприятными (внешний прямо-угольииу условиями для плодовой мушки. В 1927 г. вред, нанесенный апельсинам, был значительно больше (Боденхаймер, 1938). В Климаграммы «температура — осадки» для прибрежных районов штата Джорджия (/) на уровне моря и северной Джорджии (//) на высоте 600—900 м. В первой случае выражены влажный и сухой сезоны, что характерно для субтропического климата, а во втором сезонные различия в количестве осадков не так выражены, как различия в температуре. Растительность климатического климакса в прибрежном местообитании — широколиственный вечнозеленый лес, а в более северной области — листопадный лес умеренной зоны.[ ...]