Скорость роста популяции в естественных местообитаниях будет зависеть от климатических изменений, от снабжения пищей и от того, ограничено ли размножение определенным временем года и др., что должно учитываться при составлении моделей или их усовершенствовании.[ ...]
Скорость роста популяции — изменение численности популяции за единицу времени. Скорость роста популяции может быть положительной, нулевой и отрицательной. Она зависит от показателей рождаемости, смертности и миграции (вселения — иммиграции и выселения — эмиграции). Увеличение (прибыль) численности происходит в результате рождаемости и иммиграции особей, а уменьшение (убыль) численности — в результате смертности и эмиграции особей.[ ...]
Общая скорость роста популяции в отсутствие лимитирующего влияния среды (г) зависит от возрастного состава и вклада в репродукцию различных возрастных групп. Таким образом, вид может характеризоваться несколькими величинами г в зависимости от структуры популяции. Когда устанавливается стационарное и стабильное распределение возрастов, специфическую скорость роста называют показателем потенциального роста популяции или Гтах- Часто эту максимальную величину г называют иначе — биотический или репродуктивный потенциал. Разность между гтах, или биотическим потенциалом, и фактической скоростью роста в данных лабораторных или полевых условиях используют как меру сопротивления среды, которая характеризует сумму всех лимитирующих факторов среды, препятствующих реализации биотического потенциала.[ ...]
На уровне популяции абиотические факторы воздействуют на такие параметры, как рождаемость, смертность, средняя продолжительность жизни особи, скорость роста популяции и ее размеры, нередко являясь важнейшими причинами, определяющими характер динамики численности популяции и пространственное распределение особей в ней. Популяция может приспосабливаться к изменениям абиотических факторов, во-первых, меняя характер своего пространственного распределения и, во-вторых, путем адаптивной эволюции.[ ...]
Поскольку популяция изменчива, нас интересует не только ее величина и состав в каждый данный момент, но также и то, как она изменяется. Зная скорость изменения популяции, можно судить о многих ее важных особенностях. Скорость можно определить, разделив величину изменения на период времени, за который оно произошло; скорость, выраженная таким образом, будет характеризовать быстроту, с какой что-то изменяется во времени. За» означает «деленное на». Например, скорость роста популяции — это число организмов, на которое она увеличивается за некоторое время; ее получают путем деления величины прироста популяции на протекший промежуток времени. Средние скорости изменения популяций принято выражать в виде ДЛ /М где N — размер популяции (или другой существенный показатель), а t — время. Мгновенные скорости обозначают как йЫ1<И.[ ...]
Параметры роста популяции жертвы (“Prey Growth Parameters”): г - Удельная мгновенная скорость роста популяции жертвы; значения г должны быть положительными и лежать в интервале от 0 до 5. По умолчанию г= 1.[ ...]
Мгновенную скорость йЫ/сИ, а также скорость с?М/(Мс?/) нельзя рассчитать непосредственно, лишь на основании подсчетов численности; необходимо также знать характер кривой роста популяции. Вычисляют мгновенную скорость с помощью уравнений, которые будут рассмотрены в разд. 7. Мы не можем подсоединить к популяции «спидометр» » определить ее мгновенную скорость, как в автомобиле. Разумеется, приближенный расчет можно было бы сделать, проводя переписи численности популяции через очень короткие промежутки времени. При этом мы, вероятно, установили бы, что по средней величине совершенно нельзя-судить о том, как изменяется скорость роста в последовательные моменты времени. В приведенном выше примере специфическая скорость роста вычислена по отношению ,к величине популяции в начальный момент (50 простейших). Другими словами, на каждую из первоначальных 50 особей образовалось по 2 новые особи. При этом некоторые особ» могли бы разделиться дважды, тогда как другие совсем не разделились-бы, а часть особей вообще исчезла бы из популяции. Перепись численности в начальный и конечный моменты какого-то промежутка времени ничего не скажет нам о том, как же все это произошло на самом деле. Специфическая скорость роста может быть также выражена по отношению к средней величине популяции в течение данного периода времени. Пр» оценке годовой скорости роста популяций человека (в процентах) за основу обычно берут плотность в середине года. Так, скорость 1% в гол означает, что на каждые 100 человек популяции, имевшихся в середине-года, добавилось по одному новому индивидууму. Выражение скорости по отношению к числу имевшихся особей позволяет сравнивать скорости-роста популяций, сильно различающихся по величине, например населения небольшого и большого государства.[ ...]
Таким образом, скорость рождаемости, скорость смертности и скорость роста популяции могут быть средними и мгновенными. Далее по тексту подразумевается мгновенная скорость, однако слово «мгновенная» сознательно опущено, чтобы не перегружать текст.[ ...]
| Изолинии скоростей роста, зависящих от степени обеспеченности ресурсами. Каждая из этих изолиний соединяет точки, соответствующие таким количествам двух ресурсов (#1 и #2) в окружающей среде, которые понадобились бы для того, чтобы популяция росла с данной скоростью (для каждой изолинии— своей). С ростом обилия ресурсов скорость роста популяции увеличивается, и поэтому чем выше скорость роста, тем дальше от начала координат расположена соответствующая ей изолиния |  |
В этом уравнении скорость роста популяции зависит от ее численности в момент (t - Т), где Т- это время запаздывания (time lag) обратной связи, измеряемое числом поколений. Даже столь простая модель позволяет выявить основные эффекты запаздывания на динамику роста популяции.[ ...]
Описанные модели роста популяции и дифференциальные уравнения предполагают, что все организмы сходны между собой, имеют равную вероятность погибнуть и равную способность к размножению, так что скорость роста популяции в экспоненциальной фазе зависит только от ее численности и не ограничена условиями среды, которые остаются постоянными. Идеальность» всех экологических факторов в исходных условиях предопределила то, что рассматриваемые модели называют идеальными.[ ...]
Модели динамики и роста популяции. Любая популяция способна (теоретически) к неограниченному росту численности, если ее не лимитируют факторы внешней среды абиотического (прежде всего климат) и биотического (конкуренция, хищники, паразиты, болезни) происхождения. В таком случае, конечно, гипотетическом, скорость роста популяции будет определяться величиной ранее упоминавшегося биотического или репродуктивного потенциала.[ ...]
Обозначим численность популяции первого вида Ыи а численность второго — N2. Предельную плотность насыщения и максимальную врожденную скорость роста популяций обозначим соответственно Ки Кг, г и г2.[ ...]
Нужно подчеркнуть, что на скорость роста популяции и другие показатели оказывают сильное влияние сроки размножения. Давление отбора может приводить к возникновению различных адаптивных изменений в жизненном цикле; при этом может сдвинуться время начала размножения, но не изменится общее число потомков или же может измениться продукция или «размер кладки», но не будет затронуто время размножения. Эти и многие другие стороны динамики популяции можно выявить с помощью таблиц структуры популяции по продолжительности жизни особей.[ ...]
Вообще ’’эффект Олли”, т.е. увеличение скорости роста популяции при объединении отдельных особей во взаимодействующие группы (самым простым примером такого объединения служит возникновение репродуктивных пар) может приводить к возникновению нескольких нетривиальных положений равновесия. Переход популяции из одного состояния в другое может происходить как вследствие естественной эволюции системы, так и под действием случайных возмущений. Иногда с такими переходами связывают понятие ’’эластичности” сообщества. Точнее, система считается ’’эластичной”, если случайные воздействия не разрушают ее, а приводят в другое стационарное состояние. Среди равновесных точек системы могут встречаться как устойчивые, в окрестности которых система будет проводить большую часть времени, так и неустойчивые, которые связаны с границами областей притяжения устойчивых состояний.[ ...]
Ясно, что мгновенная удельная скорость роста популяции 6NIN6t максимальна (г = rmax) когда N = 0 и (К - N)tK = 1, и равна нулю при N = К и (К - N)IK= 0. Это означает, что популяция прекращает рост при достижении численности К, когда среда обитания оказывается полностью занятой.[ ...]
Модели зависимого от плотности роста, такие как логистическое уравнение, описывают процесс внутривидовой конкуренции, при котором по мере увеличения численности особей ресурсы становятся все более ограничивающим фактором, и удельная скорость роста популяции уменьшается. Модель межвидовой конкуренции Лотки-Вольтерры (Lotka, 1925; Volterra, 1926) построена на основе логистического уравнения и по существу несет в себе все его недостатки. Однако, несмотря на это, данная модель является наиболее простым и с исторической точки зрения очень важным способом анализа межводовой конкуренции. Она может помочь выявить основные факторы, определяющие исход конкурентного взаимодействия двух видов.[ ...]
| Зависимость численности популяции от времени (а) и скорости роста популяции от ее численности (б). Штриховкой обозначена область вырождения популяции |  |
Различают абсолютную и удельную скорость роста популяции.[ ...]
Если г положительно, численность популяции увеличивается экспоненциально. Если г отрицательно, численность популяции уменьшается экспоненциально. Отсюда быстрое увеличение и падение численности популяции. Скорость роста каждого организма не зависит от плотности популяции. Размеры популяции не стабилизируются Если Ы> К, скорость роста отрицательна. Если К > Ы, скорость роста положительна, то величина популяции стремится к К = ЛГ, т.е. приводится соответствие с поддерживающей емкостью среды. Когда К = ЛГ, скорость роста популяции равна нулю. Размеры популяции остаются постоянными.[ ...]
Хотя для обозначения рождаемости и скорости роста популяции могут быть использованы одинаковые обозначения, эти две величины — отнюдь не одно и то же, потому что через АЫ в каждом случае обозначают разные величины. В случае рождаемости АЫп — число новых особей, прибавившихся к популяции. Рождаемость может быть нулевой или положительной, но никогда — отрицательной. Что касается скорости роста популяции, то здесь ДМ — чистое увеличение или уменьшение популяции, которое является следствием не только рождаемости, но и смертности, выселения, вселения и т. д. Скорость роста популяции может быть отрицательной, нулевой или положительной, поскольку популяция может уменьшаться, оставаться неизменной или увеличиваться.[ ...]
При таких условиях неограниченного роста изменение численности популяции во времени выражается экспоненциальной кривой (рис. 12.5, А), описываемой уравнением Nt = N0 ■ е”, где Na—исходная численность Nt — численность в момент времени t, е — основание натуральных логарифмов1. Если численность отложить в логарифмическом масштабе, ее изменения выразятся прямой линией, наклон которой в системе координат определяется величиной г (рис. 12.5, Б). Описанная экспоненциальная модель роста популяции отражает ее потенциальные возможности размножения. Показатель мгновенной удельной скорости роста популяции г нередко определяют как репродуктивный потенциал популяции или ее биотический потенциал (R. Chapman, 1928, 1931). Экспоненциальный рост популяции возможен лишь при условии неизменного, независимого от численности значения коэффициента г.[ ...]
Кроме того, программа вычисляет чистую скорость размножения (по уравнению 9), среднее когортное время генерации (по уравнению 10), приближенное (по уравнению 11) и точное (по уравнению 12) значения удельной скорости роста популяции при данных начальных условиях.[ ...]
Воздействие экологических факторов на скорость роста популяции может довести численность популяции до стабильной (г=0) либо ее уменьшить, т.е. экспоненциальный рост замедляется или останавливается полностью и 1-образная кривая как бы останавливается и выполаживается, превращаясь в так называемую Б-образную кривую (рис. 3.3). В природе так и происходит — дальнейшее развитие популяции идет по логистической модели, что и описывается 8-образной, или логистической кривой роста популяции.[ ...]
Известно три типа зависимости численности популяции от ее плотности (рис. 7.10). При первом типе (кривая 1) скорость роста популяции уменьшается по мере увеличения плотности. Это широко распространенное явление позволяет понять, почему популяции некоторых животных относительно устойчивы. Во-первых, при увеличении плотности популяции наблюдается снижение рождаемости. Так, в популяции большой синицы при плотности меньше одной пары на 1 га на одно гнездо приходится 14 птенцов; когда же плотность достигает 18 пар на 1 га, выводок составляет менее 8 птенцов. Во-вторых, при увеличении плотности популяции меняется возраст наступления половой зрелости. Например, африканский слон в зависимости от плотности популяции может достигать половой зрелости в возрасте от 12 до 18 лет. Кроме того, этот вид при низкой плотности дает приплод 1 слоненок за 4 года, тогда как при высокой - рождаемость составляет 1 слоненок за 7 лет.[ ...]
Количественные показатели (характеристики) популяции можно разделить на статические и динамические. Статические показатели характеризуют состояние популяции на данный момент времени. Основные из них: численность, плотность, а также показатели структуры. Динамические показатели популяции отражают процессы, протекающие в популяции за определенный промежуток времени. Основные из них: рождаемость, смертность, скорость роста популяции.[ ...]
При отсутствии лимитирующих факторов среды удельная скорость роста равна величине г, которая характеризует свойства самой популяции и называется удельной (врожденной) скоростью роста популяции или биотическим потенциалом вида.[ ...]
Первые члены правых частей системы (9.10) характеризуют скорость роста популяций при отсутствии ограничивающих факторов. Вторые члены учитывают те изменения в скоростях, которце вызываются ограниченностью корма.[ ...]
Репродукционный (репродуктивный) потенциал способность популяции к увеличению численности, константа скорости роста популяции.[ ...]
В точках пересечения кривых выедания и пополнения чистая скорость роста популяции жертвы равна нулю (выедание равно пополнению). Каждая из этих точек характеризуется величинами плотности жертвы и хищника, и эти пары значений плотности, располагаясь на изоклине жертвы, характеризуют, таким образом, объединенные популяции. По этим точкам была построена изоклина для жертвы, приведенная на нижнем рисунке (и на рис. 10.7): ее форма характерна для самоограничивающей-ся популяции жертвы и приблизительно линейного функционального ответа. Стрелками на нижнем рисунке показано направление изменения численности жертвы.[ ...]
Динамические показатели характеризуют процессы, протекающие в популяции за какой-то промежуток (интервал) времени. Основными динамическими показателями (характеристиками) популяций являются рождаемость, смертность и скорость роста популяций.[ ...]
Конкуренция может привести к устойчивому равновесию.— Природные популяции нельзя охарактеризовать с помощью только одного параметра, такого, как предельная плотность насыщения.— Наибольшая скорость роста популяции наблюдается при промежуточных значениях плотности. Рост численности популяции при низкой плотности следует Б-образной кривой.[ ...]
Максимальная рождаемость — это теоретический верхний предел, который популяция могла бы достичь в идеальных условиях. Несмотря на трудности при практическом определении этого показателя, он представляет интерес по следующим причинам. Во-первых, максимальная рождаемость служит критерием для сопоставления с реальной рождаемостью. Например, показатель рождаемости в популяции птиц — 4 птенца в год будет иметь реальный смысл, если известен верхний предел, до которого она могла бы увеличиваться в менее лимитирующих условиях. Во-вторых, так как максимальная рождаемость — величина постоянная для данной популяции, этот показатель можно использовать для математических расчетов и прогнозирования скорости роста популяции.[ ...]
Б-образная (сигмоидная, логистическая) кривая отражает логистический тип роста, зависящего от плотности популяции, при котором скорость роста популяции снижается по мере роста численности (плотности). Скорость роста снижается вплоть до нуля при достижении предельной численности.[ ...]
Кроме того, в верхнем поле каждого графика можно увидеть вычисленные моделью значения чистой скорости воспроизводства (RQ), среднего когортного времени генерации (G), а также точной (г) и приближенной (InR0/G) величины удельной мгновенной скорости роста популяции при данных начальных условиях.[ ...]
В верхней части экрана программа выводит величины исходных параметров, а также вычисленные значения скорости роста популяции (R), средней рождаемости (Л), доли популяций, вымерших к данному поколению (Fraction Extinct), и числа прошедших поколений (Generation).[ ...]
Первое и третье решения являются устойчивыми, второе — нет. В зависимости от начальных значений численности популяции, популяция либо вырождается (если начальная численность меньше п), либо при начальной численности, большей п, развивается и достигает максимального значения, равного к. На рис. 17.5 показаны зависимость численности популяции от времени и зависимость скорости роста популяции от численности популяции [115].[ ...]
Если пренебречь показателями иммиграции и эмиграции особей (их почти нет у растений), то можно оценить мгновенную скорость роста популяции, т. е. баланс между рождаемостью и численностью за единицу времени. У стабильных популяций мгновенная скорость равна нулю, у растущих — является положительным числом, у вымирающих — отрицательным.[ ...]
Равновесие между поглощением и возобновлением ресурса. Точка К — единственная точка на ИЧСР (это не чистая скорость роста популяции), в которой концентрация ресурса ие изменяется (скорости поглощения и возобновления равны и направлены в противоположные стороны). Более подробные объяснения см. в тексте.[ ...]
| Результаты моделирования сообществ по Хьюстону. А. Ожидаемая зависимость между разнообразием и частотой сокращения популяций. Б. Ожидаемая зависимость между разнообразием и скоростью роста популяций в неравновесных системах при низких и средних частотах сокращений численности. В. Обобщенный график зависимости между скоростью конкурентного вытеснения (эквивалент скорости роста популяций), частотой сокращения популяций и видовым разнообразием. Разнообразие изменяется по оси, перпендикулярной плоскости страницы, и представлено в виде замкнутых кривых (по Huston, 1979) | 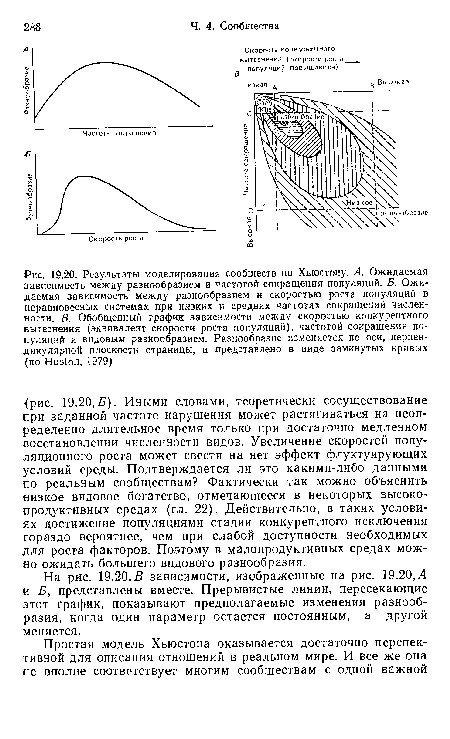 |
На печеночной двуустке (Fasciola hepatica) можно наблюдать сходный эффект скученности, а кроме того, две другие зависимости. С увеличением плотности популяции доля паразитов, достигающих половозрелости, снижается, а что еще важнее — стадия, на которой образуются яйца, наступает значительно позже. Это является непосредственной причиной снижения потенциальной скорости роста популяции двуустки.[ ...]
Ресурс может быть определен как вещество, организм или иной материальный объект, который потребляет, или иным образом использует организм, что приводит к росту удельной скорости роста его популяции при увеличении доступности ресурса. Если вид потребляет единственный ограниченный ресурс, его популяция может в конечном счете достигнуть равновесия, при котором скорость роста популяции равна скорости ее убыли, а скорость пополнения ресурса в среде равна скорости его потребления. В состоянии равновесия, когда 6NiN6t = 0 = 6Rf6t, прирост популяции равен ее убыли, а скорость поступления ресурса равна скорости его потребления. Это равновесие может быть достигнуто только при одной специфической концентрации ограниченного ресурса, обозначаемой как R . Таким образом, R - это концентрация ресурса, необходимая популяции вида для обеспечения прироста, уравновешивающего ее убыль. Одновременно это такая концентрация, до которой уровень ресурса может быть снижен потребляющей его популяцией при достижении равновесия.[ ...]
И еще одно замечание. Обратите внимание на то, что слово «вредный» ни разу не употреблялось при описании отрицательных взаимодействий. Конкуренция и хищничество снижают скорость роста популяции, подвергающейся воздействию, но этот эффект не обязательно вреден, если рассматривать его с точки зрения выживания популяции на протяжении длительного Бремени или с точки зрения ее эволюции. Отрицательные взаимодействия могут ускорять естественный отбор, приводя к возникновению новой адаптации. Как мы уже видели, хищники и паразиты часто полезны для популяций, не имеющих механизмов саморегуляции, для предотвращения перенаселения, следствием которого могло бы быть самоуничтожение (см. разд. 8).[ ...]
Из рисунка видно, что в среде, нейтральной в отношении размера, скороспелость гораздо выгоднее отсрочки, а моноцикличность— полицикличности. У скороспелых организмов время генерации короче, поэтому скорость роста популяции выше.[ ...]
Из рисунка видно, что в среде, нейтральной в отношении размера, скороспелость гораздо выгоднее отсрочки, а моноцикличность— полицикличности. У скороспелых организмов время генерации короче, поэтому скорость роста популяции выше.[ ...]
Вопрос о том, насколько законы экологии можно переносить на взаимоотношения человека с окружающей средой, остается открытым, так как человек отличается от всех других видов. Например, у большинства видов скорость роста популяции уменьшается с увеличением ее плотности; у человека, наоборот, рост населения в этом случае ускоряется. Стало быть, некоторые регулирующие механизмы природы отсутствуют у человека, и это может служить дополнительным поводом для технологического оптимизма у одних, а для экологических пессимистов свидетельствовать об опасности такой катастрофы, которая невозможна ни для одного иного вида.[ ...]
С другой стороны, развитие — это постепенная дифференци-ровка частей организма, позволяющая ему осуществлять различные функции (например, размножаться) на разных стадиях жизненного цикла. Во многих случаях рост и развитие происходят одновременно. Однако это два совершенно отдельных процесса. Одной и той же стадии онтогенеза может соответствовать широкий диапазон размеров, а экземпляры одинакового размера могут различаться по уровню развития. Быстрое развитие может быть выгодным, поскольку приближает начало размножения, сокращает время генерации и тем самым увеличивает скорость роста популяции (гл. 4). С другой стороны, если организму в течение жизни приходится переживать период крайне неблагоприятных условий, бывает выгодной остановка в развитии (т. е. стадия покоя или диапауза) (гл. 5).[ ...]
В противоположность этому два других австралийских эколога — Андреварта и Берч — смотрят на зависящие от плотности процессы в целом как на второстепенные и не играющие у некоторых видов никакой роли в определении численности популяций. В отличие от Николсона их работы в большей степени касались борьбы с насекомыми-вредителями в естественных условиях. Точка зрения Андреварты и Берча (Andrewartha, Birch, 1954) может быть кратко изложена следующим образом: «Численность животных в природной популяции ограничивается тремя способами: (а) недостатком материальных ресурсов, таких, как пища, места сооружения гнезд и т. д.; (б) недоступностью этих материальных ресурсов из-за ограниченных способностей животных расселяться и искать их; (в) ограниченным периодом времени, в течение которого величина г (скорость роста популяции) имеет положительное значение. Видимо, чаще всего встречается именно последний вариант, а реже всего — первый. Колебания величины г могут быть вызваны погодой, хищниками или любым другим компонентом среды, влияющим на скорость роста популяции».[ ...]
Тем не менее на рис. 6.8 во всех случаях наблюдается колоколообразная кривая. Форма этой кривой отражает общую природу зависимых от плотности изменений рождаемости и смертности всякий раз, когда возникает внутривидовая конкуренция.[ ...]