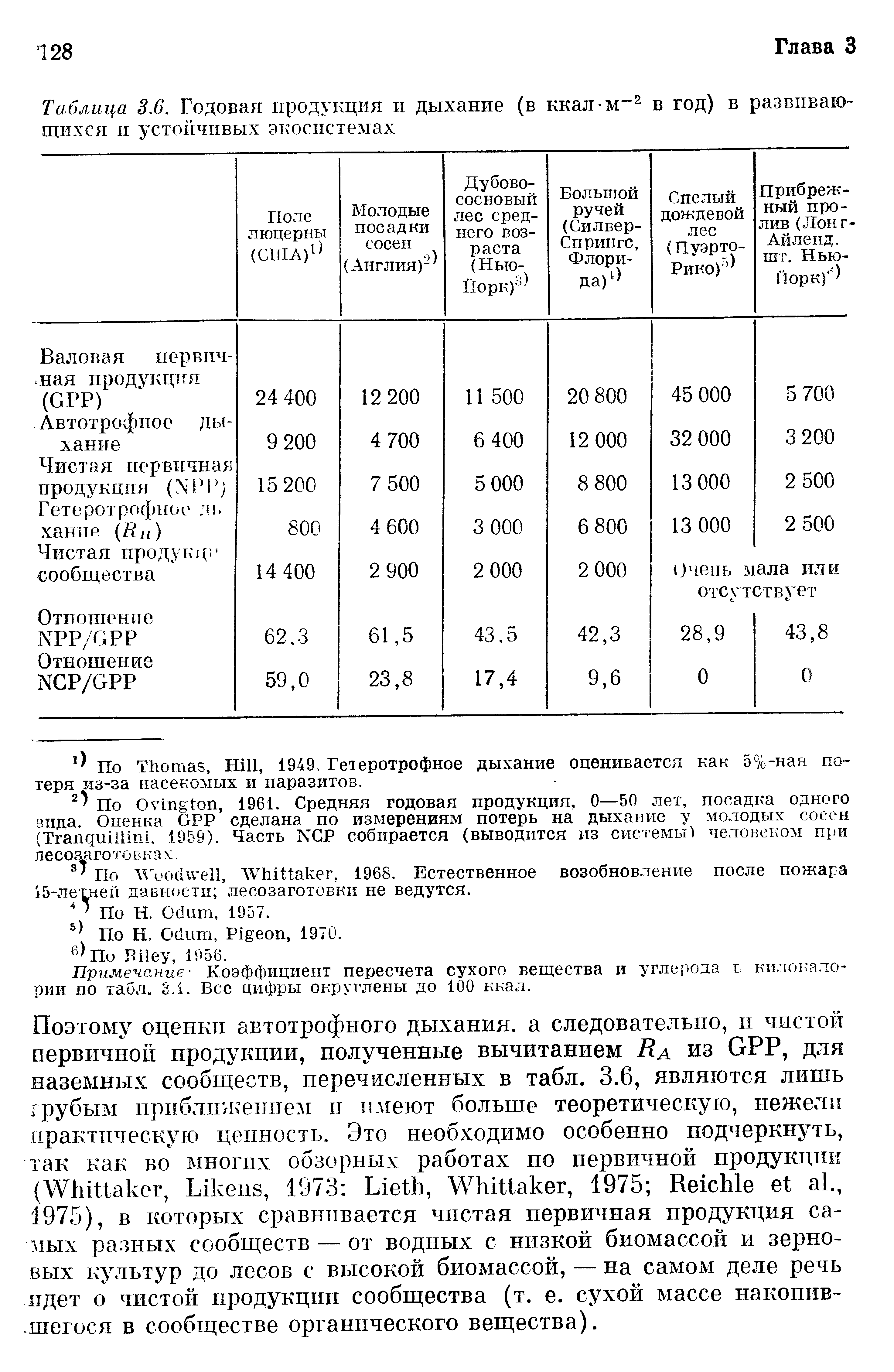
Рассматривая общую концепцию энергетических субсидий, надо сделать еще одно замечание. Фактор, который при одних условиях среды или при одном уровне поступлений увеличивает продуктивность, при других условиях среды или другом уровне поступлений может способствовать утечкам энергии, уменьшая продуктивность. Слишком много хорошего так же вредит системе, как и слишком мало. Так, усиленная эвапотранспирация в сухом климате может приводить к перерасходу энергии, а во влажном дает дополнительную энергию (Н. Odum, Pigeon, 1970). Экосистемы проточных вод, например ручей в Флориде, данные по которому включены в табл. 3.6, обычно более продуктивны, чем экосистемы стоячих вод, но не в том случае, если поток воды слишком разрушителен или нерегулярен. Ровная смена приливов и отливов на соленых маршах, в заросших манграми эстуариях или на коралловых рифах весьма способствует высокой продуктивности этих сообществ, но на северных скалистых побережьях, страдающих зимой от льда, а летом от жары, приливно-отливные течения могут отнимать много энергии у сообщества. Заболоченные леса вдоль рек, регулярно заливаемые в течение периода покоя в конце зимы и в начале весны, имеют гораздо большую продуктивность, чем леса, залитые постоянно или заливаемые надолго во время вегетативного периода (Е. Odum, 1978).~~В табл. 3.6 приведены примеры различных экосистем — от используемых человеком быстро растущих до зрелых, находящихся в стационарных состояниях. Расположив их именно в таком порядке, легко заметить некоторые важные закономерности в соотношении валовой первичной продукции (GPP), чистой первичной продукции (NPP) и чистой продукции сообщества (NCP). Системы с быстрым ростом, или «цветущие» (т. е. такие, в которых на короткое время развивается высокая продукция), например поле люцерны, обычно характеризуются высокой чистой первичной продукцией, и если они защищены от консументов, то и высокой чистой продукцией сообщества. Уменьшение гетеротрофного дыхания может быть либо результатом действия развившихся в процессе эволюции механизмов самозащиты (таких, как природные системные инсектициды или целлюлозные структуры у растений), либо следствием поступления энергии извне. В сообществах, находящихся в стационарном состоянии, валовая первичная продукция обычно полностью расходуется на автотрофное (Ra) и гетеротрофное (Rh) дыхание, так что к концу годового цикла чистая продукция сообщества очень невелика или же ее совсем не остается. Кроме того, для поддержания сообществ с большой биомассой или «урожаями на корню» типа дождевого леса необходим большой объем автотрофного дыхания, поэтому в таких сообществах отношение NPP/GPP мало (табл. 3.6). На самом деле в таких экосистемах, как лес, при измерениях трудно разделить автотрофное и гетеротрофное дыхание. Так, потребление кислорода или выделение СО2 стволом большого дерева или его корневой системой примерно поровну складывается из дыхания ассоциированных с деревом микроорганизмов (многие из которых полезны для дерева) и дыхания его Живых тканей.~~Поэтому оценки автотрофного дыхания, а следовательно, и чистой первичной продукции, полученные вычитанием Ra из GPP, для наземных сообществ, перечисленных в табл. 3.6, являются лишь грубым приближением и имеют больше теоретическую, нежели практическую ценность. Это необходимо особенно подчеркнуть, так как во многих обзорных работах по первичной продукции (Whittaker, Likens, 1973: Lieth, Whittaker, 1975; Reichle et al., 1975), в которых сравнивается чистая первичная продукция самых разных сообществ — от водных с низкой биомассой и зерновых культур до лесов с высокой биомассой, — на самом деле речь идет о чистой продукции сообщества (т. е. сухой массе накопившегося в сообществе органического вещества).
Вернуться к оглавлению