| Черты жизненных циклов двух видов рогоза и особенности их местообитаний. Индекс э2/х — отношение дисперсии к средней величине— отражает изменчивость. Данные по рогозам удовлетворяют г/Я-схеме (по МсЫаидЫоп, 1975) | 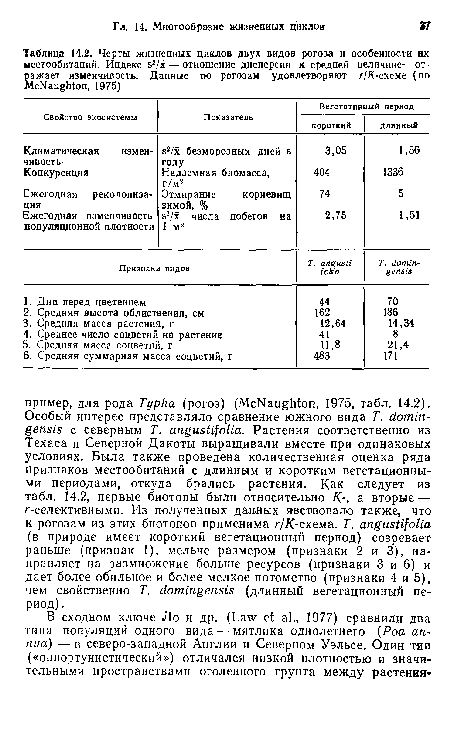 |
Черты жизненных циклов двух видов рогоза и особенности их местообитаний. Индекс э2/х — отношение дисперсии к средней величине— отражает изменчивость. Данные по рогозам удовлетворяют г/Я-схеме (по МсЫаидЫоп, 1975)
Популяционная динамика лютика ползучего на трех участках пастбища площадью 1 м2 каждый
| Популяционная динамика лютика ползучего на трех участках пастбища площадью 1 м2 каждый |  |
Типичные данные таблицы выживания для колорадского жука (в данном случае — для Меривейла за 1961—1962 гг., НагсошЧ, 1971)
| Типичные данные таблицы выживания для колорадского жука (в данном случае — для Меривейла за 1961—1962 гг., НагсошЧ, 1971) | 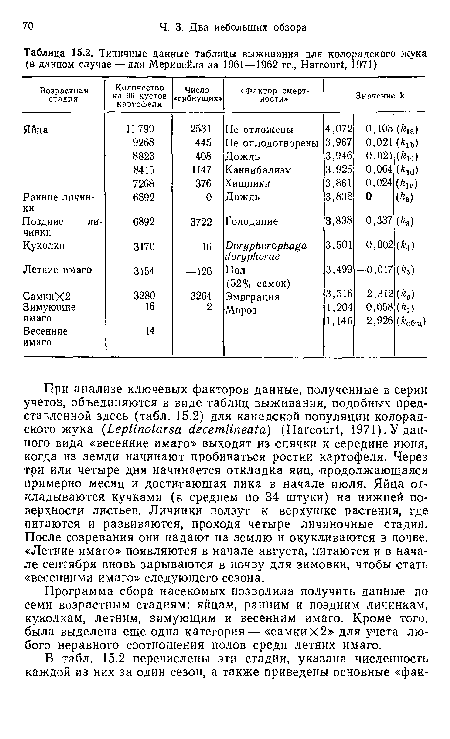 |
Показатели активности сибирского лемминга в четырех местообитаниях около Барроу. Плотности соответствуют сезонным максимумам (Brown et al, 1980)
| Показатели активности сибирского лемминга в четырех местообитаниях около Барроу. Плотности соответствуют сезонным максимумам (Brown et al, 1980) |  |
Классификация типов обычности и редкости
| Классификация типов обычности и редкости | 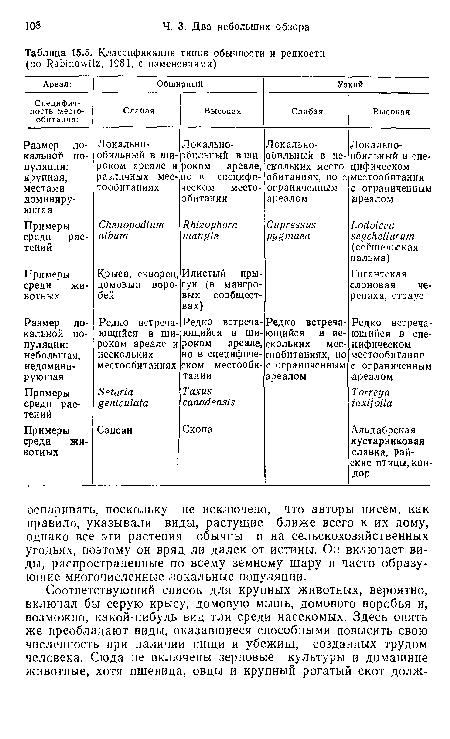 |
Некоторые характерные скорости фотосинтеза (мг СОг-дм-^ч-1) у растений одной сукцессионной последовательности. Древесные породы поздних стадий сукцессии перечислены в порядке их появления (Ваггаг, 1979)
| Некоторые характерные скорости фотосинтеза (мг СОг-дм-^ч-1) у растений одной сукцессионной последовательности. Древесные породы поздних стадий сукцессии перечислены в порядке их появления (Ваггаг, 1979) |  |
Матрица вероятностей смены через 50 лет одной древесной породы другой (Ногп, 1981)
| Матрица вероятностей смены через 50 лет одной древесной породы другой (Ногп, 1981) | 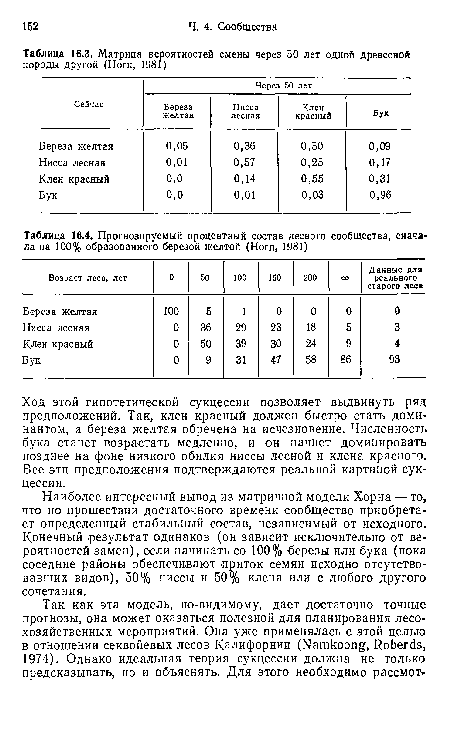 |
Прогнозируемый процентный состав лесного сообщества, снача ла на 100% образованного березой желтой (Horn, 1981)
| Прогнозируемый процентный состав лесного сообщества, снача ла на 100% образованного березой желтой (Horn, 1981) | |
Максимальные кратковременные скорости роста и эффективности фотосинтеза у различных культур на земном шаре, выраженные на единицу площади суши (Cooper, 1975; в этой работе приведены источники данных)
| Максимальные кратковременные скорости роста и эффективности фотосинтеза у различных культур на земном шаре, выраженные на единицу площади суши (Cooper, 1975; в этой работе приведены источники данных) | 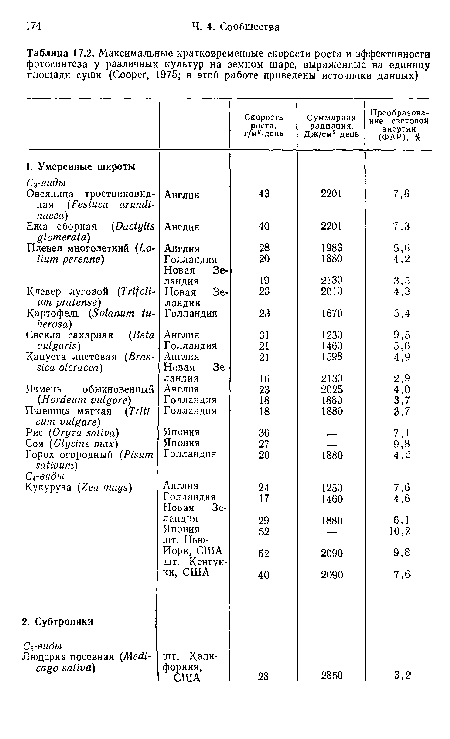 |
Модельные расчеты, иллюстрирующие влияние строения кроны на первичную продукцию, г/м2-день (Оипкап & а!., 1967)
| Модельные расчеты, иллюстрирующие влияние строения кроны на первичную продукцию, г/м2-день (Оипкап & а!., 1967) |  |
Эффективность продуцирования (Р/А X 100) у разных групп животных, расположенных в порядке ее
| Эффективность продуцирования (Р/А X 100) у разных групп животных, расположенных в порядке ее |  |
Рассчитанные величины потребления, ассимиляции, экскреции, продукции и дыхания у гетеротрофов на 100 Дж/м2 чистой годовой первичной продукции в гипотетическом степном сообществе (по Heal, MacLean, 1975)
| Рассчитанные величины потребления, ассимиляции, экскреции, продукции и дыхания у гетеротрофов на 100 Дж/м2 чистой годовой первичной продукции в гипотетическом степном сообществе (по Heal, MacLean, 1975) | 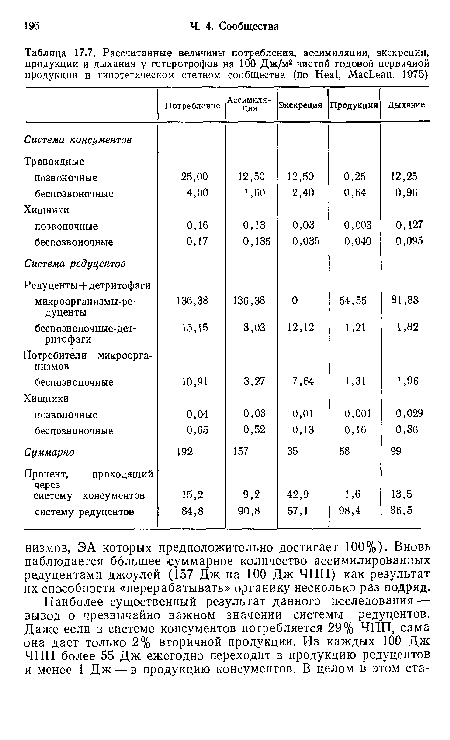 |
Основные пути поступления и потерь биогенных элементов в наземных сообществах
| Основные пути поступления и потерь биогенных элементов в наземных сообществах |  |
Основные пути поступления и потерь биогенных элементов в водных сообществах. В морских экосистемах осадки и пыль роли не играют
| Основные пути поступления и потерь биогенных элементов в водных сообществах. В морских экосистемах осадки и пыль роли не играют |  |
Схема важнейших параметров разделения ресурсов у девяти видов африканских древесных белок в вечнозеленом равнинном дождевом лесу. Горизонтальными линиями обозначено практически полное экологическое разобщение по соответствующему признаку (Emmons, 1980)
| Схема важнейших параметров разделения ресурсов у девяти видов африканских древесных белок в вечнозеленом равнинном дождевом лесу. Горизонтальными линиями обозначено практически полное экологическое разобщение по соответствующему признаку (Emmons, 1980) | 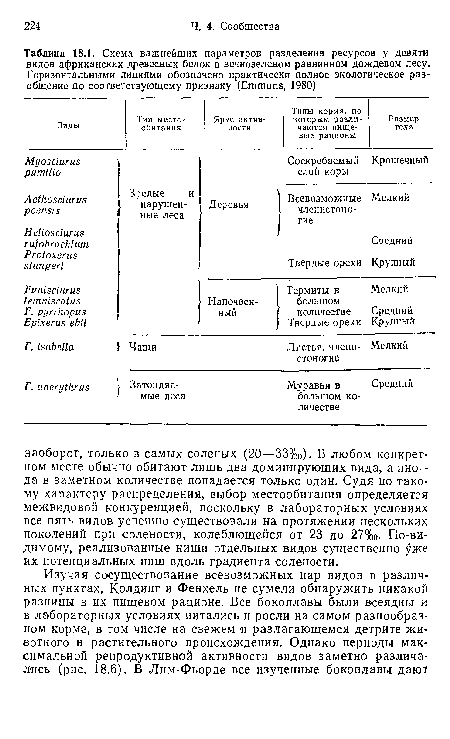 |
Сосуществующие виды растений, населяющие почвы сходного типа на канадском пастбище, проявляют тенденцию ие встречаться вместе (возможное конкурентное исключение), в то время как виды, произрастающие на разных типах почв, встречаются вместе (сосуществование при возможном разделении ниш). Отметим, что особенности почвы могут либо существовать изначально, либо возникать вследствие неодинакового поглощения питательных веществ растениями. Рассмотрены следующие виды
| Сосуществующие виды растений, населяющие почвы сходного типа на канадском пастбище, проявляют тенденцию ие встречаться вместе (возможное конкурентное исключение), в то время как виды, произрастающие на разных типах почв, встречаются вместе (сосуществование при возможном разделении ниш). Отметим, что особенности почвы могут либо существовать изначально, либо возникать вследствие неодинакового поглощения питательных веществ растениями. Рассмотрены следующие виды | 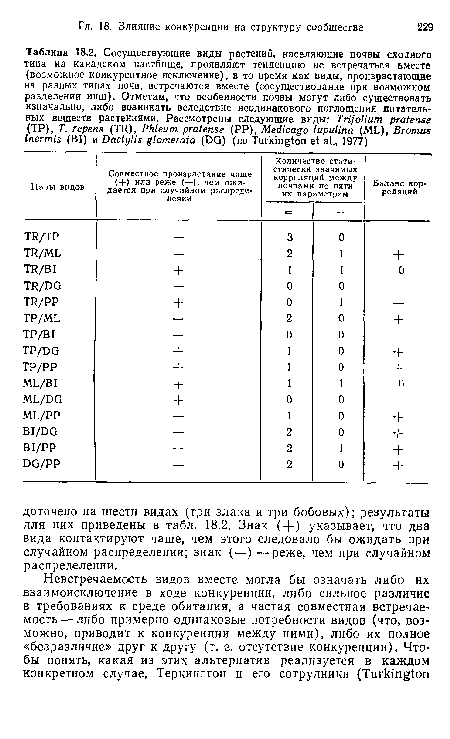 |
Проверка нуль-гипотезы о независимости локального сосуществования и перекрывания ареалов семеноядных грызуяов в пустынях Северной Америки от размеров тела. Сочетаемость считалась положительной (+) или отрицательной (—) в зависимости от того, была ли наблюдавшаяся частота совместного обитания больше или меньше случайной. Значение р отражает вероятность случайной сочетаемости. Все значеняя статистически значимы, кроме данных по пустыне Мохаве (по Bowers, 1982)
| Проверка нуль-гипотезы о независимости локального сосуществования и перекрывания ареалов семеноядных грызуяов в пустынях Северной Америки от размеров тела. Сочетаемость считалась положительной (+) или отрицательной (—) в зависимости от того, была ли наблюдавшаяся частота совместного обитания больше или меньше случайной. Значение р отражает вероятность случайной сочетаемости. Все значеняя статистически значимы, кроме данных по пустыне Мохаве (по Bowers, 1982) |  |
Число особей каждого вида, занимавшее участки, освобождавшиеся непосредственно перед учетом в результате гибели прежнего хозяина. После гибели 120 рыб трех видов свободные места были заняты 131 особью. Видовая принадлежность нового хозяина не зависела от того, кто владел участком раньше (х2=5,88, 4 степени свободы, Р>0,10)
| Число особей каждого вида, занимавшее участки, освобождавшиеся непосредственно перед учетом в результате гибели прежнего хозяина. После гибели 120 рыб трех видов свободные места были заняты 131 особью. Видовая принадлежность нового хозяина не зависела от того, кто владел участком раньше (х2=5,88, 4 степени свободы, Р>0,10) |  |
Частота встречаемости (доля участков, на которых присутствует вид) организмов, занимающих более 10% исходного пространства учетных квадратов на десяти участках нижней литорали в сентябре 1970 — мае 1979 г. (Paine, Levin, 1981)
| Частота встречаемости (доля участков, на которых присутствует вид) организмов, занимающих более 10% исходного пространства учетных квадратов на десяти участках нижней литорали в сентябре 1970 — мае 1979 г. (Paine, Levin, 1981) |  |
Основные аргументы (Elton, 1958) в пользу господствовавшей до 1970-х гг. «общепринятой» точки зрения относительно положительной корреляции сложности сообщества и его устойчивости. Все перечисленные факты согласуются с «гипотезой сложности — устойчивости», однако их можио объяснить и исходя из других обоснованных предположений, или же отвести как не поддающиеся проверке
| Основные аргументы (Elton, 1958) в пользу господствовавшей до 1970-х гг. «общепринятой» точки зрения относительно положительной корреляции сложности сообщества и его устойчивости. Все перечисленные факты согласуются с «гипотезой сложности — устойчивости», однако их можио объяснить и исходя из других обоснованных предположений, или же отвести как не поддающиеся проверке |  |
Основные структурные характеристики 40 пищевых сетей наземных, пресноводных и морских местообитаний. В столбце «трофическая структура» указано, сколько видов организмов занимает тот или иной трофический уровень, начиная с продуцентов и кончая верховными хищниками (например, формула 2-4-1-1-1 означает, что два таксона находятся на первом трофическом уровне, четыре — на втором и по одному на каждом из трех последующих). Виды, питающиеся организмами более одного трофического уровня, учитываются на самом высшем из занимаемых ими уровней. 5 — видовое богатство, С — сьязность, £ •— среднее число звеньев для всех пищевых цепей сообщества, или средняя длина максимальной пищевой цепи (по Впапё, 1983)
| Основные структурные характеристики 40 пищевых сетей наземных, пресноводных и морских местообитаний. В столбце «трофическая структура» указано, сколько видов организмов занимает тот или иной трофический уровень, начиная с продуцентов и кончая верховными хищниками (например, формула 2-4-1-1-1 означает, что два таксона находятся на первом трофическом уровне, четыре — на втором и по одному на каждом из трех последующих). Виды, питающиеся организмами более одного трофического уровня, учитываются на самом высшем из занимаемых ими уровней. 5 — видовое богатство, С — сьязность, £ •— среднее число звеньев для всех пищевых цепей сообщества, или средняя длина максимальной пищевой цепи (по Впапё, 1983) | 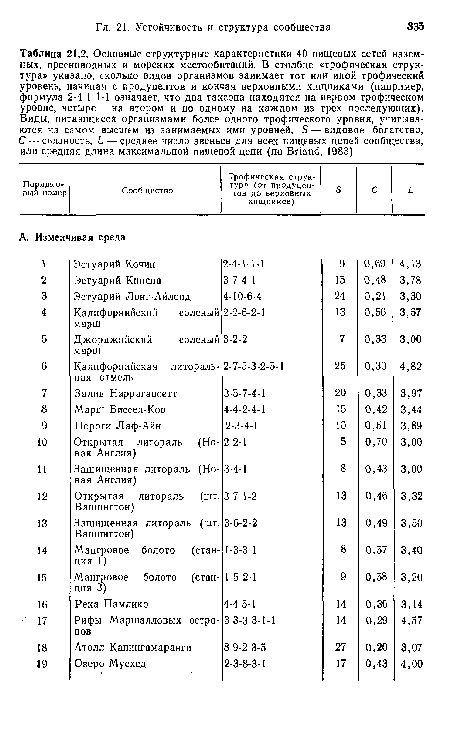 |
Влияние (А) удобрения на видовое богатство (5), равномерность распределения (Я/ 1п 5) и разнообразие (Н) растительности двух полей; (Б) стравливания бизонами на видовое разнообразие растительности двух участков (МсКаи§М
| Влияние (А) удобрения на видовое богатство (5), равномерность распределения (Я/ 1п 5) и разнообразие (Н) растительности двух полей; (Б) стравливания бизонами на видовое разнообразие растительности двух участков (МсКаи§М | 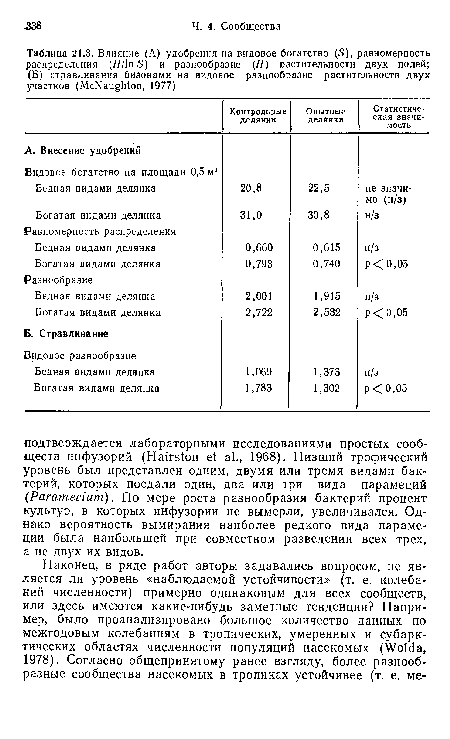 |