На рис. 6.4 представлена схема основных взаимодействий между фитопланктоном, зоопланктоном, питательными веществами, а также’ физическими и гидродинамическими характеристиками водоема. Подробное описание такой полной экосистемы со всеми взаимодействиями элементов указанной схемы будет приведенр далее в п. 6.4. Здесь же сконцентрируем наше внимание на двух планктонных членах. На первом этапе они рассматриваются отдельно, а затем исследуется их взаимодействие (в виде выедания). Для задач рационального водопользования последнее иногда (но не всегда) может рассматриваться как необязательный элемент общей схемы взаимодействий, позволяющий внести ясность относительно конкретных типов фитопланктона, формирующих фитоценоз водоема. Кстати сказать, более подробную информацию относительно разновидностей фитопланктона можно найти во многих гидробиологических публикациях, в том числе, например, [83, 595].[ ...]
Современные биологические работы, осуществляемые в рамках обсуждаемой в книге проблемы качества воды, например [439], оказываются все более и более ориентированными на исследование вопроса о роли видового разнообразия фитоценоза водоема.[ ...]
Временная зависимость в этом подходе заложена в члене, описывающем рост скорости вымирания фитопланктона за счет усиливающегося выедания зоопланктоном.[ ...]
Несколько более сложная система уравнений, учитывающих также и вариации по глубине, была проанализирована в работе Рили и др. [446]. Однако в этом подходе было принято упрощающее предположение о том, что система находится в устойчивом состоянии. Это обстоятельство ограничило применение разработанной модели лишь для летнего периода года [512]. Стил в своих модельных построениях использовал два объемных сегмента и сохранил члены, содержащие производные по времени. Однако он не посчитал нужным учесть временную изменчивость как температуры, так и приходящей солнечной радиации. Вывод всех обсуждавшихся до сих пор систем уравнений основывался на концепции сохранения массы. Основной переменной являлась масса первичной продукции, однако в рассмотрение включались также и массы зоопланктона и питательных веществ. Одним из основных различий между упомянутыми моделями, равно как и любыми другими планктонными моделями, являются заложенные в расчеты константы скоростей. Опубликованная в 1949 г. работа Монода [352] может служить канвой для анализа скоростей роста и вымирания фитопланктона.[ ...]
При этом экспоненциальный рост или спад биомассы фитопланктона зависит от знака разности (Рф—б). Задача определения знака может быть решена как в полевых условиях, так и в условиях эксперимента. Однако в общем случае скорости Рф и <3 зависят от концентраций питательных веществ Ы, уровня облученности водоема солнечным светом и наличия хищников более высокого «уровня».[ ...]
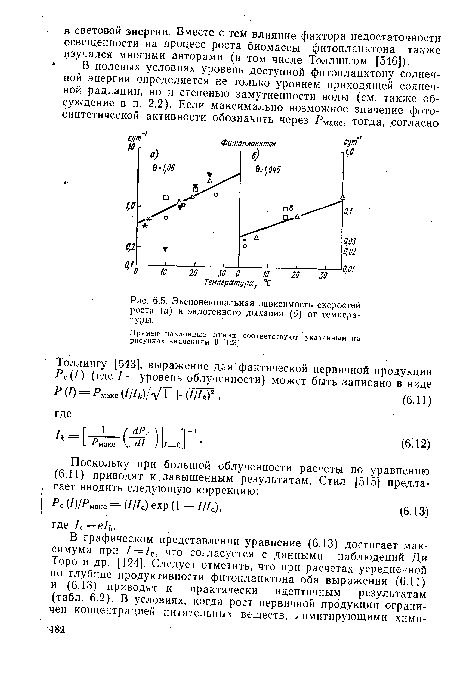
На рис. 6.5 приведены примеры (в основу которых положены данные Ди Торо и др. [124]) почти экспоненциальных скоростей роста (и дыхания) в функции температуры.[ ...]
Прямые наклонные линии соответствуют указанным на рисунках значениям 0 [123].[ ...]
Теория Монода применима для условий неизменности значений параметров, входящих в модель. В то же время было обнаружено, что при малых д имеет место более быстрое усвоение питательного элемента, т. е. Ум ос <7-1. Однако следует признать, что на практике эта разница не имеет серьезного значения [121].[ ...]
В рамках этой модели учитываются также и сезонные вариации I.[ ...]
Этот новый компонент должен быть в обязательном порядке включен в уравнение баланса массы.[ ...]
Рисунки к данной главе:
| Схема влияния внешних факторов на взаимодействующие системы фито- и зоопланктона, а также питательных веществ (биогенов) [124]. |
![Схема влияния внешних факторов на взаимодействующие системы фито- и зоопланктона, а также питательных веществ (биогенов) [124].](/static/pngsmall/901285264.png) |
| Экспоненциальная зависимость скоростей , роста (а) и эндогенного дыхания (б) от температуры. |
 |
| Схематическое представление трех вариантов усвоения питательных веществ [539]. |
![Схематическое представление трех вариантов усвоения питательных веществ [539].](/static/pngsmall/901285274.png) |