Последовательные стадии развития можно рассматривать как процесс, при котором в различные критические точки времени и пространства происходит переключение на альтернативные пути дальнейшего развития. Это переключение может наблюдаться на клеточном уровне, например, когда две дочерние клетки, возникающие в результате неэквивалентного деления, дифференцируются по-разному; она может также происходить при дифференцировке органов или даже апекса побега как целого, например при переходе от вегетативной фазы развития к цветению. Далее мы уже видели, что если орган, такой, как зачаток листа, прошел определенную стадию развития, то он необратимо «детерминируется» как лист (в отличие от почки) п обычно не может превратиться ни в одну другую структуру (с. 53—54).[ ...]
Такое последовательное вступание различных частей развивающегося организма на специфические пути дифференцировки называют «канализацией развития». Так, развитие зародыша включает канализацию по нескольким основным путям разви-тия, в результате чего появляются области корня и побега. С появлением организованного апекса побега возникает основа для заложения различных органов (лист, почка, стебель), которая продолжается во время всей последующей вегетативной фазы развития.[ ...]
Коль скоро группа клеток вступила на какой-то путь развития, она обычно следует по этому «нормальному» пути до полного его завершения, и крайне редко клетки возвращаются к более ранней стадии развития или переходят на какой-либо другой путь. Так, листовые примордии не станут почками или стеблями, хотя иногда при формировании цветка могут возникать, аномалии развития, например возврат к вегетативной верхушке, по такие случаи сравнительно редки, поэтому считают, что на определенных критических стадиях те или иные части организма становятся «детерминированными» в отношении их дальнейшей дифференцировки. Мы уже приводили пример такой детерминации при развитии листовых примордиев (рис. 2.12).[ ...]
Мы не располагаем такими же примерами для растении, но данные о существовании детерминации растительных клеток и тканей имеются. Наиболее ярко детерминация проявляется у апексов побега. Само предположение о детерминации мернсте-матических клеток может показаться парадоксальным, поскольку именно эти клетки обычно считают недифференцированными и не вступившими на какой-либо путь развития. Однако мы уже. рассматривали два или три примера детерминации апикальных меристем стебля, а именно яровизацию и смену фаз у древесных растений. Как мы уже видели, при яровизации зародышей ржи происходят внутренние изменения в потенциях развивающихся клеток меристемы стебля, так что они становятся способными к цветению. При полной яровизации зародышей ржи все клетки, произошедшие из апикальной меристемы, становятся яровизированными, и это состояние, очевидно, передается от •одного поколения клеток к другому без «разбавления» (с. 361). Хотя яровизированное состояние очень стабильно, новые зародыши, образованные яровизированным растением, уже не яровизированы. Следовательно, при нормальном гаметогенезе должна происходить деяровизация.[ ...]
Еще одним примером детерминации верхушечных меристем является «смена фаз» у древесных растений (с. 381). Смена фаз, несомненно, связана с внутренними изменениями меристс-матических клеток, так как если выращивать в стерильной культуре на одинаковой питательной среде каллусы из тканей ювенильных и взрослых побегов плюща, то эти культуры различаются между собой. Каллус из ювенильных побегов растет быстрее и образует больше корней, чем каллус из взрослых побегов. Очевидно, здесь мы имеем дело с двумя сравнительно • стабильными альтернативными состояниями детерминации, которые не связаны с генетическими изменениями (поскольку на взрослых побегах образуются семена, из которых развиваются ювенильные проростки), но которые могут передаваться от одного поколения клеток к другому без утраты детерминации.[ ...]
Мы мало знаем о том, с какими изменениями на молекулярном уровне связана детерминация. Представляется вероятным, что мы здесь имеем дело со стабильными состояниями генной экспрессии, которые не изменяются даже после клеточного деления. Трудно представить себе, каким образом различия в активности генов могут сохраняться в ходе репликации ДНК и Последующего цитокинеза, но передача дифференцированного состояния через повторные клеточные деления хорошо установлена для животных клеток.[ ...]
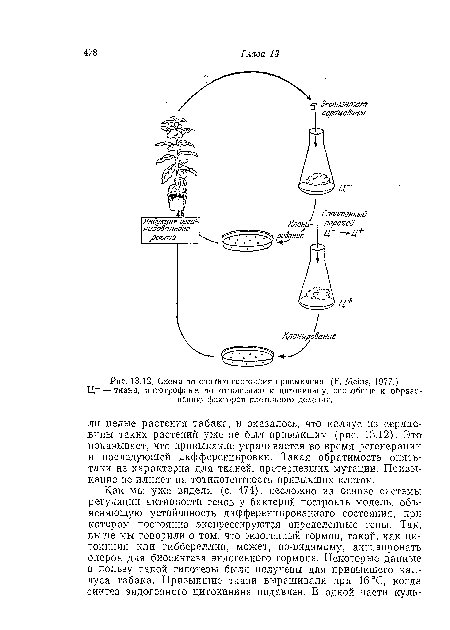
Несмотря на скудость наших знаний относительно молекулярной основы детерминации, мы можем все же сказать, что между явлениями детерминации и явлением «привыкания» растительных клеток в стерильной культуре (с. 237) существует близкая аналогия. Известно, что в среду для культивирования каллуса из тканей сердцевины табака обычно необходимо добавлять экзогенные ИУК и цитокиннн. Следовательно, клетки сердцевины табака, по-видпмому, или неспособны к синтезу ауксина и цитокинииа и поэтому нуждаются в их добавке, или, возможно, они очень быстро катаболизируют эти ростовые вещества. Вместе с тем исходно гетеротрофные по отношению к ауксинам и цитокининам культуры могут в ходе культивирования стать «привыкшими» и сами удовлетворять свои потребности в ростовых веществах того или иного типа. Таким образом, культура сердцевины табака, по-видимому, может существовать в устойчивых альтернативных состояниях, очень напоминая этим детерминированные ткани. Важно, что клоны «привыкших» тканей, полученные из одной клетки, все еще остаются привыкшими; следовательно, возникшие изменения являются свойствами индивидуальных клеток.[ ...]
Ц- — ткани, автотрофные по отношению к цитокинииу, способные к образованию факторов клеточного деления.[ ...]
Если мы правы в своем заключении, что дедифференцировка и перепрограммирование в значительной мере снимают предшествующее состояние избирательного маскирования генома, то это разрешает кажущееся противоречие между концепциями детерминации и тотипотеитности. В свете этого предположения, по-видимому, не существует никакого противоречия между предполагаемым состоянием детерминации апексов стебля и корня и образованием придаточных корней и почек из взрослых тканей соответственно стеблей и корней.[ ...]
Рисунки к данной главе:
| Схема по снятию состояния привыкания. (F. Meins, 1977.) |
 |