Как мы видели, для зацветания короткодневных растений они не обязательно должны постоянно находиться в условиях короткого дня. После определенного числа благоприятных фото-периодических циклов КД-растеиия зацветут, даже если их перенести в условия длинного дня. Так, например, ХапШит в конечном итоге зацветает в ответ и на один КД-Цикл, даже если после него следует значительное число ДД-циклов. Следовательно, некоторые виды проявляют выраженную фотопериоди-ческую «устойчивость», иными словами, растения становятся «индуцированными» после получения КД.[ ...]
Для объяснения устойчивости были предложены две теории. Согласно предположению М. X. Чайлахяна, под влиянием благоприятных условий в листе накапливается гормон цветения, который расходуется постепенно в течение длительного периода даже при неблагоприятных условиях. Другой советский исследователь Б. С. Мошков предположил, что под влиянием благоприятной длины дня в метаболизме листа происходят стойкие изменения, в результате чего он продолжает активно синтезировать гормон цветения даже после перенесения в условия неблагоприятного фотопериода. Имеющиеся в нашем распоряжении данные говорят в пользу теории Мошкова. В самом деле, кажется маловероятным, чтобы в описанном выше эксперименте Лона в листе, получившем КД- ВОЗдействие и после этого находившемся в условиях ДД в течение 4 нед, гормон цветения ■оставался в количестве, все еще достаточном для индукции цветения вегетирующего растения, на которое этот лист был привит.[ ...]
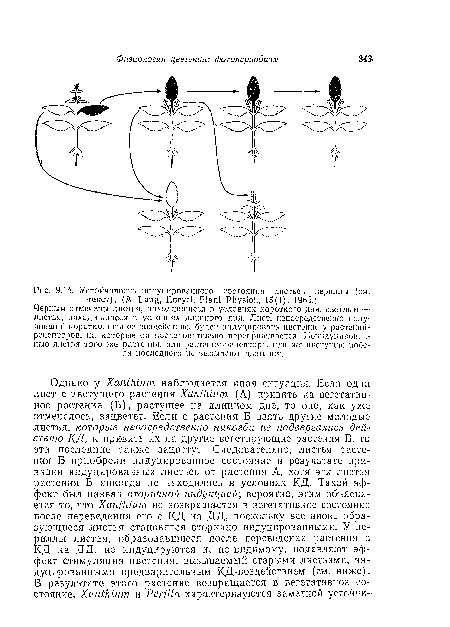
Установлено, что у растений периллы состояние индукции строго локализовано в пределах растения и даже в пределах отдельных листьев. Так, если одну пару листьев периллы подвергнуть действию ряда КД-Циклов, а остальную часть растения содержать в условиях ДД, то оказывается, что только листья, непосредственно получившие КД-воздействие, способны индуцировать цветение у растений, на которые они привиты (рис. 9.14). В других экспериментах, проведенных Лона, в условиях КД находились лишь половины каждого листа, а другие половины получали ДД. Затем листья разделяли продольно и половинки прививали по отдельности на вегетативные растения-рецепторы. Цветение вызывали лишь те половинки, которые непосредственно подвергались действию КД.[ ...]
Рисунки к данной главе:
| Устойчивость индуцированного состояния листьев периллы (см. |
 |