Тем не менее зависимость доза—эффект в фототропизме намного сложнее, чем кажется с первого взгляда. Так, в экспериментах на этиолированных колеоптилях было установлено, что с увеличением количества раздражения изгиб по направлению к источнику света увеличивается, но до определенной пороговой величины (приблизительно ОД Дж м 2 световой энергии), превышение которой ведет к снижению ответной реакции до некоторого начального значения, а иногда «положительная реакция» может даже перейти в «отрицательную» (т. е. изгиб <в сторону, противоположную источнику света). При дальнейшем увеличении количества раздражения реакция может снова стать положительной (рис. 7.4). Как было установлено, закон количества раздражения Бансена—Роско справедлив только для первой положительной и первой отрицательной реакций.[ ...]
Колеоптили других зерновых характеризовались такой же зависимостью, за’ исключением того, что не всегда имели четко выраженную первую отрицательную реакцию.[ ...]
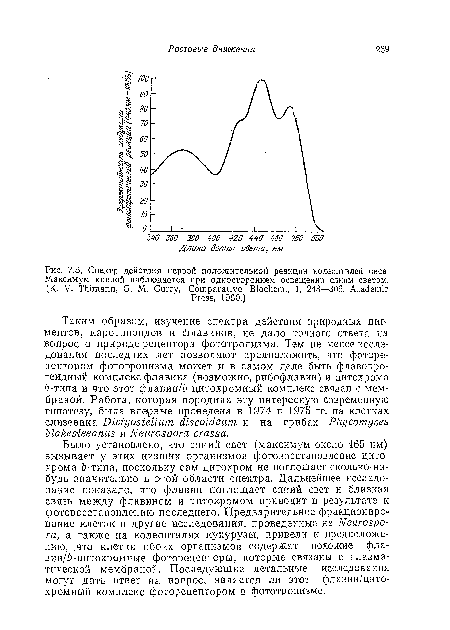
Точные измерения показали, что спектр действия положительного фототропизма у спораигиеносцев фикомицетов почти идентичен таковому для колеоптилей овса и других зерновых. Следовательно, подобный фоторецептор, видимо, существует и у большинства организмов. Такие спектры действия наводят на мысль, что фоторецептором в фототропизме является желтый пигмент. Наиболее вероятно, что им может быть либо каротиио-ид, либо флавин. Спектры поглощения (З-каротииа и рибофлавина показаны на рис. 7.6; хотя оба пигмента имеют максимумы поглощения в синей области, ни один из спектров поглощения полностью не совпадает со спектром действия фототропизма (рис. 7.5).[ ...]
Фитохром — фоторецептор, поглощающий красный и дальний красный свет и принимающий участие в многочисленных явлениях фотомо рфогенеза растений (гл. 8), — по-видимому, иепринимает непосредственного участия в фототропизме. Однако известно, что освещение красным светом (не направленным) влияет на чувствительность органов растения к направленному синему свету, причем эффект красного света может быть изменен на противоположный при освещении дальним красным. Полученные результаты свидетельствуют о том, что фитохром представляет собой пигмент, который служит посредником эффектов красного света в фототропизме, одиако неясно, каким образом фитохром связан с системой фототропизма.[ ...]
Согласно трем основным выдвинутым гипотезам, передача раздражения в фототропизме обусловлена неодинаковыми уровнями ауксина в освещенной и затененной сторонах органа.[ ...]
Согласно этой гипотезе, синтез ауксина на освещенной стороне верхушки колеоптиля подавляется.[ ...]
Заметим, что если гипотезы 2 и 3 справедливы, то одностороннее освещение верхушки колеоптиля должно приводить к уменьшению общего количества содержащегося в верхушке ауксина. Гипотеза 1 предполагает, что одностороннее раздражение верхушки светом не должно отраоюаться на общем содержании ауксина.[ ...]
Теперь в первую очередь рассмотрим три гипотезы, основанные на латеральной асимметрии уровня ауксина. Гипотеза прямой светоростовой реакции будет рассмотрена ниже (7.6.3).[ ...]
Рисунки к данной главе:

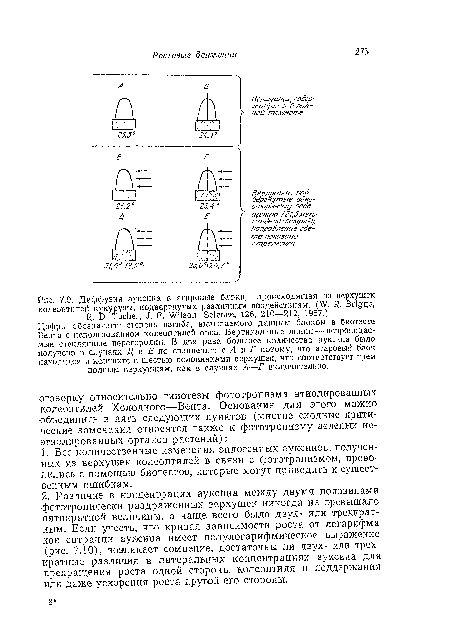
| Диффузия ауксина в агаровые блоки, происходящая из верхушек колеоптилей кукурузы, подвергнутых различным воздействиям. (W. R, Briggs, Я. D. Tocher, J. F. Wilson, Science, 126, 210—212, 1957.) |
 |