Нам хорошо известно, что ауксины, а, возможно, также и другие гормоны роста, такие, как гиббереллииы и этилен, могут изменять механические свойства стенок растительных клеток, однако до сих пор неизвестно, каким образом достигаются эти эффекты. Поскольку большая часть данных по связанным с ростом изменениям в клеточной стенке получена для ауксинов, в последующем изложении мы в основном и будем рассматривать рост, индуцированный этим гормоном.[ ...]
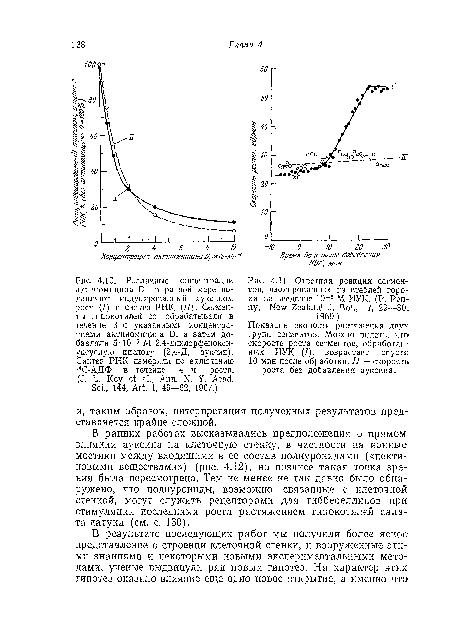
В настоящее время известно, что хотя для роста клеток растяжением требуется длительный синтез белка и РНК (рис. 4.10), а также активное дыхание, тем не менее если ткани стебля или колеоптиля обрабатывать ауксином, то скорость роста увеличится после очень небольшого лаг-периода, всего лишь порядка нескольких минут (рис. 4.11). Поэтому кажется маловероятным, чтобы рост ускорялся благодаря изменениям в скорости транскрипции или трансляции. Скорее ауксин влияет на какую-то «предсуществующую» систему. Также хорошо установлено, что в ответ на действие ауксина происходят значительные изменения в полисахаридах клеточной стенки; и в самом деле трудно себе представить, чтобы без таких изменений могли измениться механические свойства клеточной стенки. Поскольку в данном случае происходят разрыв и образование новых ковалентных связей, очевидно, что в этих изменениях участвуют ферменты. Исследователи давно пытались выяснить, какие именно изменения происходят в полисахаридах клеточной стенки и коррелируют ли эти изменения с различиями в активности ферментов внутри стенки.[ ...]
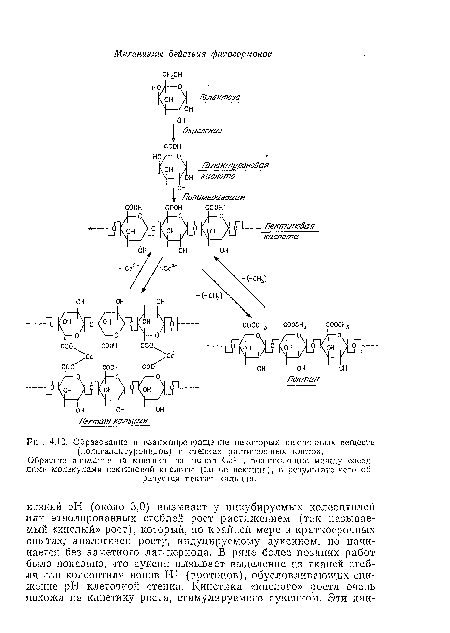
В ранних работах высказывались предположения о прямом влиянии ауксина на клеточную стенку, в частности на ионные мостики между входящими в ее состав полиуронидами («пектиновыми веществами») (рис. 4.12), но позднее такая точка зрения была пересмотрена. Тем не менее не так давно было обнаружено, что полиурониды, возможно связанные с клеточной стенкой, могут служить рецепторами для гиббереллинов при стимуляции последними роста растяжением гипокотилей салата-латука (см. с. 130).[ ...]
Обратите вннмаппе па мостики пз попов СаУ -, возникающие между соседними молекулами пектиновой кислоты (но не пектина), в результате чего образуется псктат кальция.[ ...]
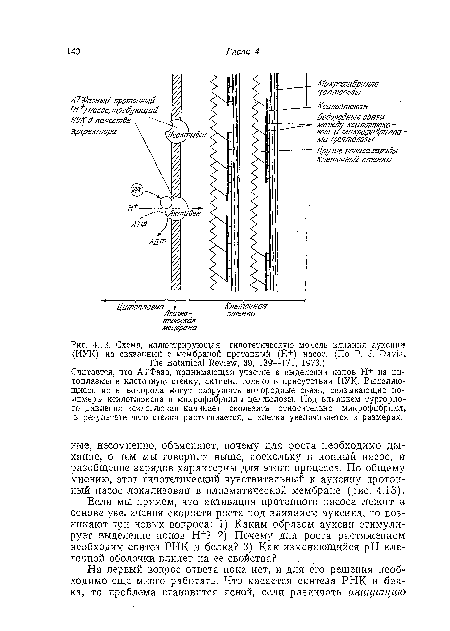
Считается, что АТФаза, принимающая участие в выделении ионов Н 1 из цитоплазмы в клеточную стенку, активна только в присутствии ИУК. Выделяющиеся ионы водорода могут разрушать водородные связи, связывающие полимеры ксилоглюкана и микрофибриллы целлюлозы. Под влиянием тургорно-го давления ксилоглюкан начинает скользить относительно микрофибрилл, в результате чего стенка растягивается, а клетка увеличивается в размерах.[ ...]
Несомненно, что для нормального длительного роста растяжением необходим непрерывный синтез белка, даже если краткосрочные (с лаг-периодом порядка 10 мин или меньше) ростовые ответы на гормон наблюдаются и без синтеза новых РНК и белков. В случае регуляции роста растяжением ауксином очевидно, что при быстром ростовом ответе происходит выделение И+ в оболочку, а снижение pH или непосредственно ослабляет какие-то межмолекулярные связи, или благоприятствует активности определенных ферментов, разрыхляющих оболочку.[ ...]
Влияние других гормонов на увеличение размеров клеток пока не изучено так подробно, как влияние ауксинов. Латентный период для стимуляции роста клеток гиббереллинами, по-видимому, значительно длиннее (не менее 30 мни по сравнению с 10 мин для ауксина), и еще не вполне ясно, происходит ли при этом выделение Н+ растущими клетками. Вместе с тем этилен изменяет скорость роста менее чем за 5 мин после помещения тканей в его атмосферу.[ ...]
Итак, механизмы, лежащие в основе роста клеток растений растяжением, постепенно выясняются, и роль ауксина в этом процессе становится более понятной. Однако не следует забывать, что, хотя увеличение размера клеток во многих отношениях является наиболее характерной ответной реакцией на действие ауксинов, эта реакция не единственная и, очевидно, даже не всегда первая. Так, ауксин может индуцировать быстрое увеличение интенсивности дыхания и скорости движения протоплазмы. Кроме того, ряд ответных реакций на действие ауксина не связан с немедленной вакуолизацией клеток (например, деление клеток камбия, образование корней, коррелятивное ипгии бирование пазушных почек). Другими словами, хотя ауксин может вызывать быстрое разрыхление клеточной стенки, это отт шодь не единственный способ его действия.[ ...]
Рисунки к данной главе:
| Ответная реакция сегментов, изолированных из стеблей гороха, на введение 10“5 М ИУК. (P. Penny, New Zealand J. Bot., 7, 29—30,. |
 |
| Образование и взаимопревращение некоторых пектиновых веществ (полигалактуропидов) в стенках растительных клеток. |
 |
| Схема, иллюстрирующая гипотетическую модель влияния аукснпа (ИУК) на связанный с мембраной протонный (Н+) насос. (По P. J. Davis, The Botanical Review, 39, 139—171, 1973.) |
 |